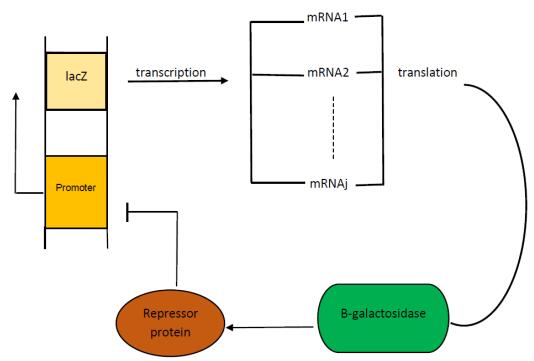
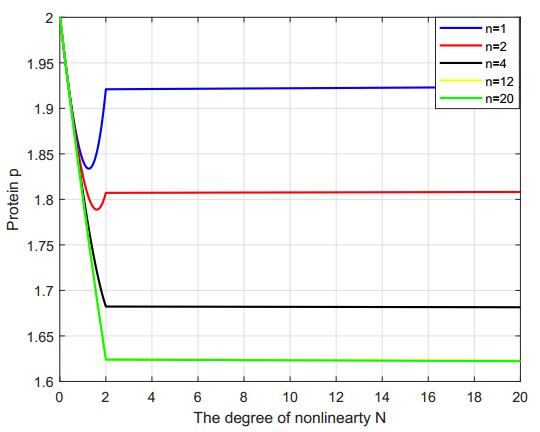
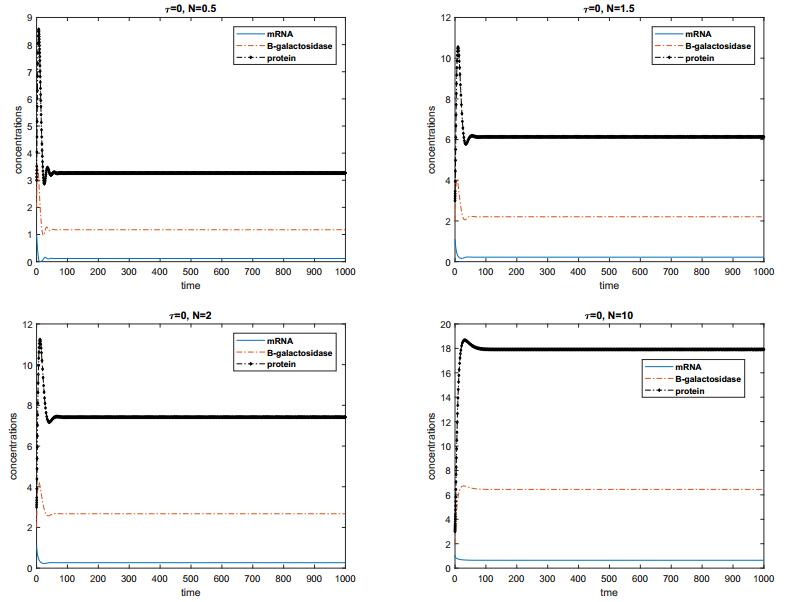
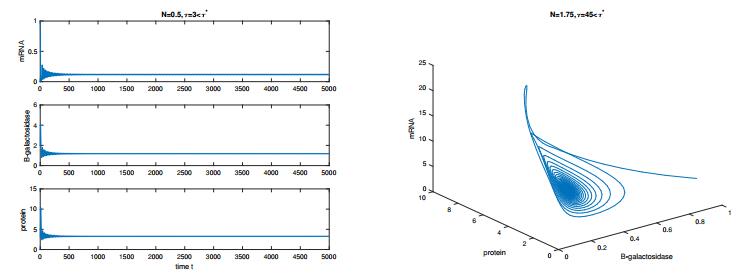
In this paper, we construct a discrete time delay Lac operon model with nonlinear degradation rate for mRNA, resulting from the interaction among several identical mRNA pieces. By taking a discrete time delay as bifurcation parameter, we investigate the nonlinear dynamical behaviour arising from the model, using mathematical tools such as stability and bifurcation theory. Firstly, we discuss the existence and uniqueness of the equilibrium for this system and investigate the effect of discrete delay on its dynamical behaviour. Absence or limited delay causes the system to have a stable equilibrium, which changes into a Hopf point producing oscillations if time delay is increased. These sustained oscillation are shown to be present only if the nonlinear degradation rate for mRNA satisfies specific conditions. The direction of the Hopf bifurcation giving rise to such oscillations is also determined, via the use of the so-called multiple time scales technique. Finally, numerical simulations are shown to validate and expand the theoretical analysis. Overall, our findings suggest that the degree of nonlinearity of the model can be used as a control parameter for the stabilisation of the system.
Citation: Zenab Alrikaby, Xia Liu, Tonghua Zhang, Federico Frascoli. Stability and Hopf bifurcation analysis for a Lac operon model with nonlinear degradation rate and time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1729-1749. doi: 10.3934/mbe.2019083
In this paper, we construct a discrete time delay Lac operon model with nonlinear degradation rate for mRNA, resulting from the interaction among several identical mRNA pieces. By taking a discrete time delay as bifurcation parameter, we investigate the nonlinear dynamical behaviour arising from the model, using mathematical tools such as stability and bifurcation theory. Firstly, we discuss the existence and uniqueness of the equilibrium for this system and investigate the effect of discrete delay on its dynamical behaviour. Absence or limited delay causes the system to have a stable equilibrium, which changes into a Hopf point producing oscillations if time delay is increased. These sustained oscillation are shown to be present only if the nonlinear degradation rate for mRNA satisfies specific conditions. The direction of the Hopf bifurcation giving rise to such oscillations is also determined, via the use of the so-called multiple time scales technique. Finally, numerical simulations are shown to validate and expand the theoretical analysis. Overall, our findings suggest that the degree of nonlinearity of the model can be used as a control parameter for the stabilisation of the system.

| [1] | B. Goodwin, Oscillatory behaviour in enzymatic control process, Adv. Enzyme Regul., 3 (1965), 425–438. |
| [2] | S. Busenberg and J. Mahaffy, Interaction of spatial diffusion and delays in models of genetic control by repression, J. Math. Biol., 22 (1985), 313–333. |
| [3] | J. Mahaffy and C. Pao, Models of genetic control by repression with time delays and spatial effects, J. Math. Biol., 20 (1984), 39–57. |
| [4] | M. Monk, Oscillatory expression of Hes1, p53, and NF-k B driven by transcription time delays, Curr. Biol., 13 (2003), 1409–1413. |
| [5] | S. Nikolov, J. Vera, V. Kotev, et al., Dynamic properties of a delayed protein cross talk model, BioSystems, 91 (2008), 483–500. |
| [6] | Y. Song, X. Cao and T. Zhang, Bistability and delay-induced stability switches in a cancer network with the regulation of microRNA, Commun. Nonlinear Sci. Numer. Simul., 54 (2018), 302–319. |
| [7] | N. Yildirim, M. Santillan and D. Horike, Dynamics and bi-stability in a reduced model of the lac operon, Chaos, 14 (2004), 279–291. |
| [8] | N. Yildirim and M. Mackey, Feedback regulation in the Lactose operon: a Mathematical modeling study and comparison with experimental data, Biophys. J., 84 (2003), 2841–2851. |
| [9] | A. Verdugo, Dynamics of gene networks with time delays, Ph.D thesis, Cornell university, USA, 2009. |
| [10] | K. Wang, L. Wang, Z. Teng, et al., Stability and bifurcation of genetic regulatory networks with delays, Neurocomputing, 73 (2010), 2882–2892. |
| [11] | H. Zang, T. Zhang and Y. Zhang, Bifurcation analysis of a mathematical model for genetic regulatory network with time delays, Appl. Math. Comput., 260 (2015), 204–226. |
| [12] | M. B. Elowitz and S. Leibler, A synthetic oscillatory network of transcription regulators, Nature, 403 (2000), 335–338. |
| [13] | D. Liu, X. Chang, Z. Liu, et al., Bistability and Oscillations in Gene Regulation Mediated by Small Noncoding RNAs, PLoS ONE, 6 (2011), e17029. |
| [14] | H. Liu, F. Yan and Z. Liu, Oscillatory dynamics in a gene regulatory network mediated by small RNA with time delay, Nonlinear Dyn., 76 (2014), 147–159. |
| [15] | C. Sears and W. Garrett, Microbes, Microbiota and Colon Cancer, Nonlinear Dyn., 15 (2014), 317–328. |
| [16] | Q. Xie, T. Wanga, C. Zeng, et al., Predicting fluctuations-caused regime shifts in a time delayed dynamics of an invading species, Physica A, 493 (2018), 69–83. |
| [17] | C. Zeng, Q. Xie, T. Wang, et al., Stochastic ecological kinetics of regime shifts in a time-delayed lake eutrophication ecosystem, Ecosphere, 8 (2017), 1–34. |
| [18] | J. Zeng, C. Zeng, Q. Xie, et al., Different delays-induced regime shifts in a stochastic insect outbreak dynamics, Physica A, 462 (2016), 1273–1285. |
| [19] | C. Zeng and H. Wangb, Noise and large time delay: Accelerated catastrophic regime shifts in ecosystems, Ecol. Model., 233 (2012), 52–58. |
| [20] | C. Zeng, Q. Han, T. Yang, et al., Noise- and delay-induced regime shifts in an ecological system of vegetation, J. Stat. Mech., 10 (2013), P10017. |
| [21] | Q. Han, T. Yang, C. Zeng, et al., Impact of time delays on stochastic resonance in an ecological system describing vegetation, Physica A, 408 (2014), 96–105. |
| [22] | B. D. Aguda, Y. Kim, M. G. Piper-Hunter, et al., MicroRNA regulation of a cancer network: Consequences of the feedback loops involving miR-17-92, E2F, and Myc, PNAS, 105 (2008), 19678–19683. |
| [23] | S. Bernard, J. Belair and M. C. Macky, Sufficient conditions for stability of linear differential equations with distributed delay, Discrete Cont. Dyn-S B, 1 (2001), 233–256. |
| [24] | G. A. Calin, C Sevignani, C. D. Dumitru, et al., Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers, Proc. Natl. Acad. Sci. USA, 101 (2004), 2999–3004. |
| [25] | Y. Gao, B. Feng, S. Han, et al., The Roles of MicroRNA-141 in Human Cancers: From Diagnosis to Treatment, Cell Physiol. Biochem., 38 (2016), 427–448. |
| [26] | Y. Gao, B. Feng, L. Lu, et al., MiRNAs and E2F3: a complex network of reciprocal regulations in human cancers, Oncotarget, 8 (2017), 60624–60639. |
| [27] | I.V. Makunin, M. Pheasant, C. Simons, et al., Orthologous MicroRNA Genes Are Located in Cancer Associated Genomic Regions in Human and Mouse, PLoS ONE, 11 (2007), e1133. |
| [28] | F. Yan, H. Liu, J. Hao, et al., Dynamical Behaviors of Rb-E2F Pathway Including Negative Feedback Loops Involving miR449, PLoS ONE, 7 (2009), e43908. |
| [29] | Y. Peng and T. Zhang, Stability and Hopf bifurcation analysis of a gene expression model with diffusion and time delay, Abstr. Appl. Anal., 2014 (2014), 1–9. |
| [30] | H. Wang and J. Wang, Hopf-pitchfork bifurcation in a two-neuron system with discrete and distributed delays, Math, Meth, Appl, Sci., 38 (2015), 4967–4981. |
| [31] | A. Verdugo and R. Rand, Hopf bifurcation analysis for a model of genetic regulatory system with delay, Commun. Nonlinear Sci. Number. Simul., 13 (2008), 235–242. |
| [32] | J. Wei and C. Yu, Hopf bifurcation analysis in a model of oscillatory gene expression with delay, the Royal Society of Edinburgh, 139 (2009), 879–895. |
| [33] | X. Cao, Y. Song and T. Zhang, Hop bifurcation and delay induce Turing instability in a diffusive lac operon model, Int. J. Bifurcat. Chaos, 26 (2016), 1650–1667. |
| [34] | J. Yu and M. Peng, Local Hopf bifurcation analysis and global existence of periodic solutions in a gene expression model with delays, Nonlinear Dyn., 86 (2016), 245–256. |
| [35] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, 3rd edition, New York: Springer, 2004. |
| [36] | A. H. Nayfe, Order reduction of retarded nonlinear systems- the method of multiple scales versus center-manifold reduction, Nonlinear Dyn., 51 (2008), 483–500. |
| [37] | T. Zhang, Y. Song and H. Zang, The stability and Hopf bifurcation analysis of a gene expression model, J. Math. Anal. Appl., 395 (2012), 103–113. |
| [38] | L. Zheng, M. Chen and Q. Nie, External noise control in inherently stochastic biological systems, J. Math. Phys., 53 (2012), 115616. |
| [39] | R. Cruz, R. Perez-Carrasco, P. Guerrero, et al., Minimum Action Path Theory Reveals the Details of Stochastic Transitions Out of Oscillatory States, Phys. Rev. Lett., 120 (2018), 128102. |
| [40] | N. Guisoni, D. Monteoliva and L. Diambra, Promoters Architecture-Based Mechanism for Noise- Induced Oscillations in a Single-Gene Circuit, PLoS ONE, 11 (2016), e0151086. |
| [41] | B. Ingalls, Mathematical Modelling in Systems Biology: An Introduction, 1st edition, Cambridge, MA, USA, 2013. |
| [42] | D. A. Potoyan and P. G. Wolynes, On the dephasing of genetic oscillators, PNAS, 6 (2014), 2391–2396. |
| [43] | F. Jacob, D. Perrin, C. Sanchez, et al., Lac operon: A Group of genes whose expression is coordinated by an operator, CR Biol., 250 (1960), 1727–1729. |
| [44] | S. Liu and L. Wang, Global stability of an HIV-1 model with distributed intracellular delays and a combination therapy, Math. Biosci. Eng., 7 (2010), 675–685. |
| [45] | M. Santilla and M. Mackey, Origin of Bistability in the lac Operon, Biophys. J., 92 (2007), 3830–3842. |
| [46] | A.Wan and X. Zou, Hopf bifurcation analysis for a model of genetic regulatory system with delay, J. Math. Anal. Appl., 365 (2009), 464–476. |
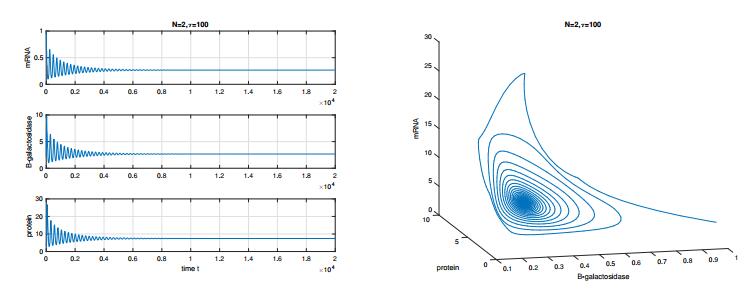
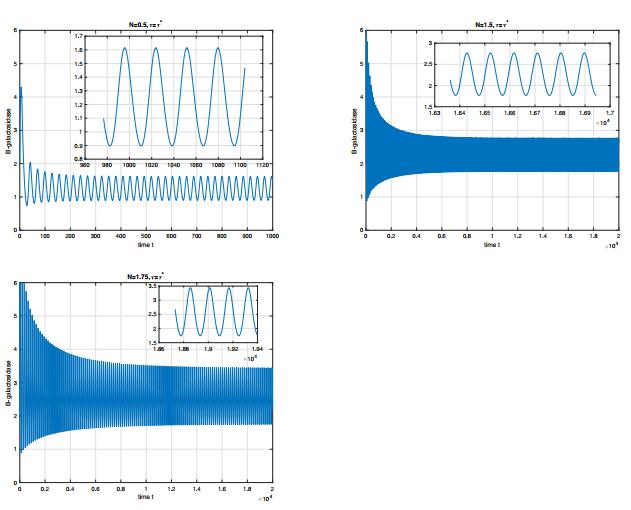
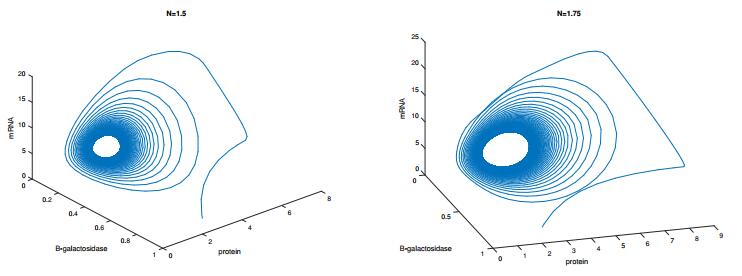
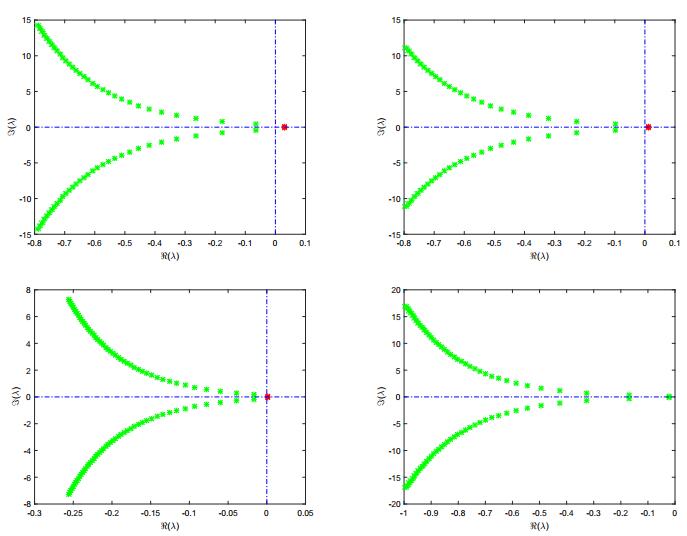
Figures(8) / Tables(1)

Zenab Alrikaby, Xia Liu, Tonghua Zhang, Federico Frascoli. Stability and Hopf bifurcation analysis for a Lac operon model with nonlinear degradation rate and time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1729-1749. doi: 10.3934/mbe.2019083







 DownLoad:
DownLoad: