Citation: Yue Liu, Wing-Cheong Lo. Analysis of spontaneous emergence of cell polarity with delayed negative feedback[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1392-1413. doi: 10.3934/mbe.2019068

| [1] | E. Bi and H. O. Park, Cell polarization and cytokinesis in budding yeast, Genetics, 191 (2012), 347–387. |
| [2] | D. M. Bryant and K. E. Mostov, From cells to organs: building polarized tissue, Nat. Rev. Mol. Cell Biol., 9 (2008), 887–901. |
| [3] | D. G. Drubin and W. J. Nelson, Origins of cell polarity, Cell, 84 (1996), 335–344. |
| [4] | M. E. Lee and V. Vasioukhin, Cell polarity and cancer–cell and tissue polarity as a non-canonical tumor suppressor, J. Cell Sci., 121 (2018), 1141–1150. |
| [5] | C. Royer and X. Lu, Epithelial cell polarity: a major gatekeeper against cancer?, Cell Death. Differ., 18 (2011), 1470–1477. |
| [6] | H. O. Park and E. Bi, Microbiology and Molecular Biology Reviews, Microbiol. Mol. Biol. Rev., 71 (2007), 48–96. |
| [7] | H. R. Bourne, D. A. Sanders and F. McCormick, The GTPase superfamily: a conserved switch for diverse cell functions, Nature, 348 (1990), 125–132. |
| [8] | S. Etienne-Manneville and A. Hall, Rho GTPases in cell biology, Nature, 420 (2002), 629–635. |
| [9] | S. J. Altschuler, S. B. Angenent, Y. Wang and L. F. Wang, On the spontaneous emergence of cell polarity, Nature, 454 (2008), 886–889. |
| [10] | B. D. Slaughter, S. E. Smith and R. Li, Symmetry breaking in the life cycle of the budding yeast, Csh. Perspect. Biol., 1 (2009), a003384. |
| [11] | A. B. Goryachev and M. Leda, Many roads to symmetry breaking: molecular mechanisms and theoretical models of yeast cell polarity, Mol. Biol. Cell, 28 (2017), 370–380. |
| [12] | J. M. Johnson, M. Jin and D. J. Lew, Symmetry breaking and the establishment of cell polarity in budding yeast, Curr. Opin. Genet. Dev., 21 (2011), 740–746. |
| [13] | S. G. Martin, Spontaneous cell polarization: Feedback control of Cdc42 GTPase breaks cellular symmetry, BioEssays, 37 (2015), 1193–1201. |
| [14] | W. C. Lo, H. O. Park and C.S. Chou, Mathematical analysis of spontaneous emergence of cell Polarity, Bull. Math. Biol., 76 (2014), 1835–1865. |
| [15] | Y. Mori, A. Jilkine and L. Edelstein-Keshet, Wave-pinning and cell polarity from a bistable reaction-diffusion system, Biophys J., 94 (2008), 3684–3697. |
| [16] | A. Rätz and M. Röger, Turing instabilities in a mathematical model for signaling networks, J. Math. Biol., 65 (2012), 1215–1244. |
| [17] | A. Rätz and M. Röger, Symmetry breaking in a bulk–surface reaction–diffusion model for signalling networks, Nonlinearity, 27 (2014), 1805–1827. |
| [18] | A. B. Goryachev and A. V. Pokhilko, Dynamics of Cdc42 network embodies a Turing-type mechanism of yeast cell polarity, FEBS Lett., 582 (2008), 1437–1443. |
| [19] | W. R. Holmes, M. A. Mata and L. Edelstein-Keshet, Local perturbation analysis: A computational tool for biophysical reaction-diffusion models, Biophys. J., 108 (2015), 230–236. |
| [20] | M. Das, T. Drake, D. J. Wiley, P. Buchwald1, D. Vavylonis and F. Verde, Oscillatory dynamics of Cdc42 GTPase in the control of polarized growth, Science, 337 (2012), 239–243. |
| [21] | A. S. Howell, M. Jin, C. F.Wu, T. R. Zyla, T. C. Elston and D. J. Lew, Negative feedback enhances robustness in the yeast polarity establishment circuit, Cell, 149 (2012), 322–333. |
| [22] | M. E. Lee, W. C. Lo, K. E. Miller, C. S. Chou and H.-O. Park, Regulation of Cdc42 polarization by the Rsr1 GTPase and Rga1, a Cdc42 GTPase-activating protein, in budding yeast, J. Cell Sci., 128 (2015), 2106–2117. |
| [23] | S. Okada, M. Leda, J. Hanna, N. S. Savage, E. Bi and A. B. Goryachev, Daughter cell identity emerges from the interplay of Cdc42, septins, and exocytosis, Dev. Cell, 26 (2013), 148–161. |
| [24] | C. F. Wu and D. J. Lew, Beyond symmetry-breaking : competition and negative feedback in GTPase regulation, Trends Cell Biol., 23 (2013), 476–483. |
| [25] | B. Xu and P. C. Bressloff, A PDE-DDE model for cell polarization in fission yeast, SIAM J. Appl. Math., 76 (2016), 1844–1870. |
| [26] | A. M. Turing, The chemical basis of morphogenesis, Bull Math. Biol., 52 (1990), 153–197. |
| [27] | W. C. Lo, M. E. Lee, M. Narayan, C. S. Chou and H. O. Park, Polarization of diploid daughter cells directed by spatial cues and GTP hydrolysis of Cdc42 in budding yeast, PLoS One, 8 (2013), 1–14. |
| [28] | H. Smith, An introduction to delay differential equations with applications to the life sciences, Springer, 2011. |
| [29] | D. Cusseddu, L. Edelstein-Keshet, J. A. Mackenzie, S. Portet and A. Madzvamuse, A coupled bulk-surface model for cell polarisation, J. Theor. Biol., (2018), 0022–5193. |
| [30] | B. Klnder, T. Freisinger, R. Wedlich-Sldner, and E. Frey, GDI-Mediated cell polarization in yeast provides precise spatial and temporal control of Cdc42 signaling, PLoS Comput. Biol., 9 (2013), 1–12. |
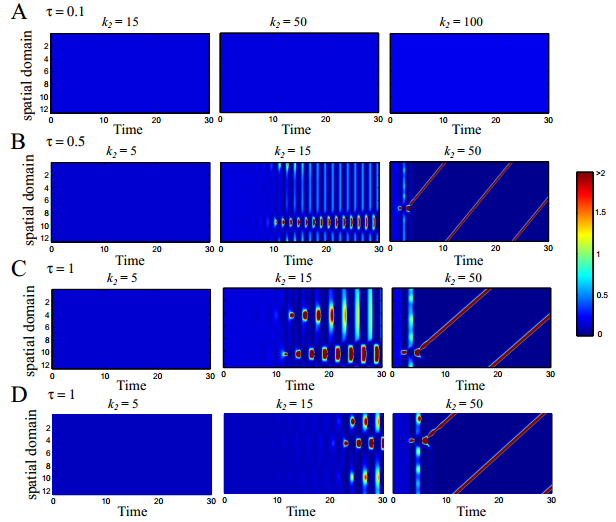
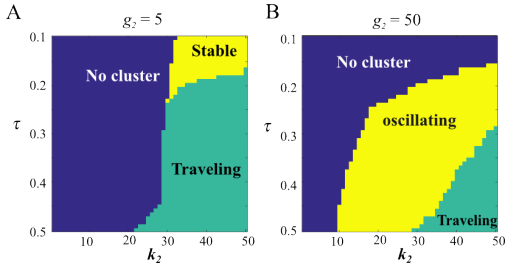
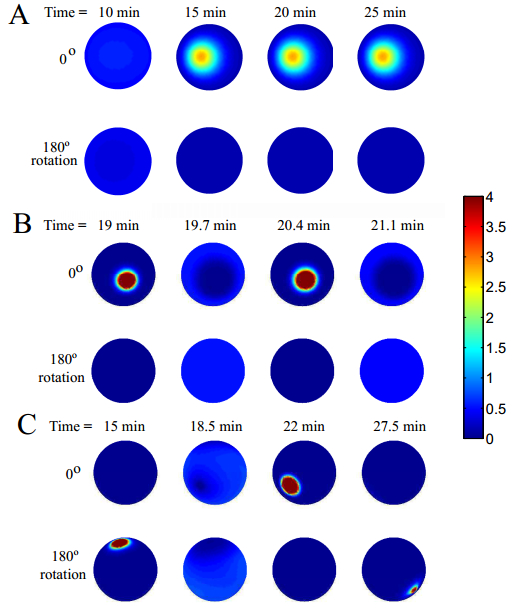
Figures(7)

Yue Liu, Wing-Cheong Lo. Analysis of spontaneous emergence of cell polarity with delayed negative feedback[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1392-1413. doi: 10.3934/mbe.2019068







 DownLoad:
DownLoad: