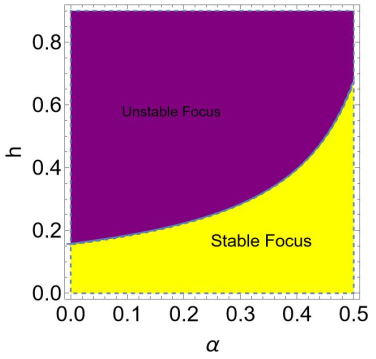
In this paper, we explore the bifurcation, chaos, and local stability of a discrete Hepatitis C virus infection model. More precisely, we studied the local stability at fixed points of a discrete Hepatitis C virus model. We proved that at a partial infection fixed point, the discrete HCV model undergoes Neimark-Sacker bifurcation, but no other local bifurcation exists at this fixed point. Moreover, it was also proved that period-doubling bifurcation does not occur at liver-free, disease-free, and total infection fixed points. Furthermore, we also examined chaos control in the understudied discrete HCV model. Finally, obtained theoretical results were confirmed numerically.
Citation: Abdul Qadeer Khan, Ayesha Yaqoob, Ateq Alsaadi. Neimark-Sacker bifurcation, chaos, and local stability of a discrete Hepatitis C virus model[J]. AIMS Mathematics, 2024, 9(11): 31985-32013. doi: 10.3934/math.20241537
In this paper, we explore the bifurcation, chaos, and local stability of a discrete Hepatitis C virus infection model. More precisely, we studied the local stability at fixed points of a discrete Hepatitis C virus model. We proved that at a partial infection fixed point, the discrete HCV model undergoes Neimark-Sacker bifurcation, but no other local bifurcation exists at this fixed point. Moreover, it was also proved that period-doubling bifurcation does not occur at liver-free, disease-free, and total infection fixed points. Furthermore, we also examined chaos control in the understudied discrete HCV model. Finally, obtained theoretical results were confirmed numerically.

| [1] |
S. Pan, S. P. Chakrabarty, Hopf bifurcation and stability switches induced by humoral immune delay in hepatitis C, Indian J. Pure Ap. Mat., 51 (2020), 1673–1695. https://doi.org/10.1007/s13226-020-0489-2 doi: 10.1007/s13226-020-0489-2

|
| [2] |
A. Mojaver, H. Kheiri, Dynamical analysis of a class of hepatitis C virus infection models with application of optimal control, Int. J. Biomath., 9 (2016), 1650038. https://doi.org/10.1142/S1793524516500388 doi: 10.1142/S1793524516500388
|
| [3] |
M. Gümüş, K. Türk, Dynamical behavior of a hepatitis B epidemic model and its NSFD scheme, J. Appl. Math. Comput., 70 (2024), 3767–3788. https://doi.org/10.1007/s12190-024-02103-6 doi: 10.1007/s12190-024-02103-6
|
| [4] |
H. Kong, G. Zhang, K. Wang, Stability and Hopf bifurcation in a virus model with self-proliferation and delayed activation of immune cells, Math. Biosci., 17 (2020), 4384–4405. https://doi.org/10.3934/mbe.2020242 doi: 10.3934/mbe.2020242
|
| [5] |
X. Jiang, J. Li, B. Li, W. Yin, L. Sun, X. Chen, Bifurcation, chaos, and circuit realisation of a new four-dimensional memristor system, Int. J. Nonlin. Sci. Num., 24 (2023), 2639–2648. https://doi.org/10.1515/ijnsns-2021-0393 doi: 10.1515/ijnsns-2021-0393
|
| [6] |
Q. Chen, B. Li, W. Yin, X. Jiang, X. Chen, Bifurcation, chaos and fixed-time synchronization of memristor cellular neural networks, Chaos Soliton. Fract., 171 (2023), 113440. https://doi.org/10.1016/j.chaos.2023.113440 doi: 10.1016/j.chaos.2023.113440
|
| [7] | Q. He, P. Xia, C. Hu, B. Li, Public information, actual intervention and inflation expectations, Transform. Bus. Econ., 21 (2022), 644. |
| [8] |
H. D. Qu, M. U. Rahman, M. Arfan, M. Salimi, S. Salahshour, A. Ahmadian, Fractal-fractional dynamical system of Typhoid disease including protection from infection, Eng. Comput., 39 (2023), 1553–1562. https://doi.org/10.1007/s00366-021-01536-y doi: 10.1007/s00366-021-01536-y
|
| [9] |
X. Zhu, P. Xia, Q. He, Z. Ni, L. Ni, Ensemble classifier design based on perturbation binary salp swarm algorithm for classification, Comput. Model. Eng. Sci., 135 (2023), 653–671. https://doi.org/10.32604/cmes.2022.022985 doi: 10.32604/cmes.2022.022985
|
| [10] |
M. Chong, M. Shahrill, L. Crossley, A. Madzvamuse, The stability analyses of the mathematical models of hepatitis C virus infection, Mod. Appl. Sci., 9 (2015), 250. https://doi.org/10.5539/mas.v9n3p250 doi: 10.5539/mas.v9n3p250
|
| [11] |
H. Dahari, A. Lo, R. M. Ribeiro, A. S. Perelson, Modeling hepatitis C virus dynamics: Liver regeneration and critical drug efficacy, J. Theor. Biol., 247 (2007), 371–381. https://doi.org/10.1016/j.jtbi.2007.03.006 doi: 10.1016/j.jtbi.2007.03.006
|
| [12] |
T. C. Reluga, H. Dahari, A. S. Perelson, Analysis of hepatitis C virus infection models with hepatocyte homeostasis, SIAM J. Appl. Math., 69 (2009), 999–1023. https://doi.org/10.1137/080714579 doi: 10.1137/080714579
|
| [13] | E. A. Grove, G. Ladas, Periodicities in nonlinear difference equations, New York: Chapman and Hall/CRC, 2004. https://doi.org/10.1201/9781420037722 |
| [14] | A. Wikan, Discrete dynamical systems with an introduction to discrete optimization problems, London, 2013. http://10.6.20.12:80/handle/123456789/30732 |
| [15] | M. R. Kulenović, G. Ladas, Dynamics of second-order rational difference equations: With open problems and conjectures, New York: Chapman and Hall/CRC, 2001. https://doi.org/10.1201/9781420035384 |
| [16] | E. Camouzis, G. Ladas, Dynamics of third-order rational difference equations with open problems and conjectures, New York: Chapman and Hall/CRC, 2007. https://doi.org/10.1201/9781584887669 |
| [17] | J. Guckenheimer, P. Holmes, Nonlinear oscillations, dynamical systems, and bifurcations of vector fields, Springer, 1983. https://doi.org/10.1007/978-1-4612-1140-2 |
| [18] | Y. A. Kuznetsov, Elements of applied bifurcation theorey, New York: Springer Science & Business Media, 2004. https://doi.org/10.1007/978-1-4757-3978-7 |
| [19] |
S. M. Rana, Chaotic dynamics and control of discrete ratio-dependent predator-prey system, Discrete Dyn. Nat. Soc., 2017 (2017), 4537450. https://doi.org/10.1155/2017/4537450 doi: 10.1155/2017/4537450
|
| [20] |
A. Rahman, D. Blackmore, Neimark-Sacker bifurcations and evidence of chaos in a discrete dynamical model of walkers, Chaos Soliton. Fract., 91 (2016), 339–349. https://doi.org/10.1016/j.chaos.2016.06.016 doi: 10.1016/j.chaos.2016.06.016
|
| [21] |
K. S. A. Basyouni, A. Q. Khan, Discrete-time predator-prey model with bifurcations and chaos, Math. Probl. Eng., 2020 (2020), 8845926. https://doi.org/10.1155/2020/8845926 doi: 10.1155/2020/8845926
|
| [22] |
H. N. Agiza, E. M. Elabbssy, H. E. Metwally, A. A. Elsadany, Chaotic dynamics of a discrete prey-predator model with Holling type II, Nonlinear Anal. Real, 10 (2009), 116–129. https://doi.org/10.1016/j.nonrwa.2007.08.029 doi: 10.1016/j.nonrwa.2007.08.029
|
| [23] |
X. Liu, D. Xiao, Complex dynamic behaviors of a discrete-time predator-prey system, Chaos Soliton. Fract., 32 (2007), 80–94. https://doi.org/10.1016/j.chaos.2005.10.081 doi: 10.1016/j.chaos.2005.10.081
|
| [24] |
P. Chakraborty, U. Ghosh, S. Sarkar, Stability and bifurcation analysis of a discrete prey-predator model with square-root functional response and optimal harvesting, J. Biol. Syst., 28 (2020), 91–110. https://doi.org/10.1142/S0218339020500047 doi: 10.1142/S0218339020500047
|
| [25] | W. Liu, D. Cai, Bifurcation, chaos analysis and control in a discrete-time predator-prey system, Adv. Differ. Equ., 2019, (2019), 11. https://doi.org/10.1186/s13662-019-1950-6 |
| [26] |
A. Q. Khan, J. Ma, D. Xiao, Bifurcations of a two-dimensional discrete time plant-herbivore system, Commun. Nonlinear Sci., 39 (2016), 185–198. https://doi.org/10.1016/j.cnsns.2016.02.037 doi: 10.1016/j.cnsns.2016.02.037
|
| [27] |
A. Q. Khan, J. Ma, D. Xiao, Global dynamics and bifurcation analysis of a host-parasitoid model with strong Allee effect, J. Biol. Dyn., 11 (2017), 121–146. https://doi.org/10.1080/17513758.2016.1254287 doi: 10.1080/17513758.2016.1254287
|
| [28] |
E. Ott, C. Grebogi, J. A. Yorke, Controlling chaos, Phys. Rev. Lett., 64 (1990), 1196. https://doi.org/10.1103/PhysRevLett.64.1196 doi: 10.1103/PhysRevLett.64.1196
|
| [29] |
D. Blackmore, A. Rahman, J. Shah, Discrete dynamical modeling and analysis of the R-S flip-flop circuit, Chaos Soliton. Fract., 42 (2009), 951–963. https://doi.org/10.1016/j.chaos.2009.02.032 doi: 10.1016/j.chaos.2009.02.032
|
| [30] |
A. Rahman, Y. Joshi, D. Blackmore, Sigma map dynamics and bifurcations, Regul. Chaotic Dyn., 22 (2017), 740–749. https://doi.org/10.1134/S1560354717060107 doi: 10.1134/S1560354717060107
|
| [31] |
A. Rahman, D. Blackmore, Interesting bifurcations in walking droplet dynamics, Commun. Nonlinear Sci., 90 (2020), 105348. https://doi.org/10.1016/j.cnsns.2020.105348 doi: 10.1016/j.cnsns.2020.105348
|
| [32] | Y. Joshi, D. Blackmore, A. Rahman, Generalized attracting horseshoes and chaotic strange attractors, arXiv Preprint, 2016, 1–8. https://doi.org/10.48550/arXiv.1611.04133 |
| [33] |
K. Murthy, I. Jordan, P. Sojitra, A. Rahman, D. Blackmore, Generalized attracting horseshoe in the Rössler attractor, Symmetry, 13 (2020), 30. https://doi.org/10.3390/sym13010030 doi: 10.3390/sym13010030
|





Figures(6)

Abdul Qadeer Khan, Ayesha Yaqoob, Ateq Alsaadi. Neimark-Sacker bifurcation, chaos, and local stability of a discrete Hepatitis C virus model[J]. AIMS Mathematics, 2024, 9(11): 31985-32013. doi: 10.3934/math.20241537







 DownLoad:
DownLoad: