Citation: Laura Hatchondo, Carole Guillevin, Mathieu Naudin, Laurence Cherfils, Alain Miranville, Rémy Guillevin. Mathematical modeling of brain metabolites variations in the circadian rhythm[J]. AIMS Mathematics, 2020, 5(1): 216-225. doi: 10.3934/math.2020013

| [1] |
B. P. Brennan, S. L. Rauch, J. E. Jensen, et al. A critical review of magnetic resonance spectroscopy studies of obsessive-compulsive disorder, Biol. Psychiat., 73 (2013), 24-31. doi: 10.1016/j.biopsych.2012.06.023

|
| [2] |
R. Guillevin, C. Menuel, J.-N. Vallée, et al. Mathematical modeling of energy metabolism and hemodynamics of WHO grade II gliomas using in vivo MR data, CR Biol., 334 (2011), 31-38. doi: 10.1016/j.crvi.2010.11.002
|
| [3] |
L. Hatchondo, N. Jaafari, N. Langbour, et al. 1H magnetic resonance spectroscopy suggests neural membrane alteration in specific regions involved in obsessive-compulsive disorder, Psychiatry Res. Neuroimaging, 269 (2017), 48-53. doi: 10.1016/j.pscychresns.2017.08.010
|
| [4] | J. Keener and J. Sneyd, Mathematical physiology, Second edition, Interdisciplinary Applied Mathematics, Springer-Verlag, New York, 2009. |
| [5] | M. Law, S. Yang, H. Wang, et al. Glioma grading: sensitivity, specificity, and predictive values of perfusion MR imaging and proton MR spectroscopic imaging compared with conventional MR imaging, Am. J. Neuroradiol., 24 (2003), 1989-1998. |
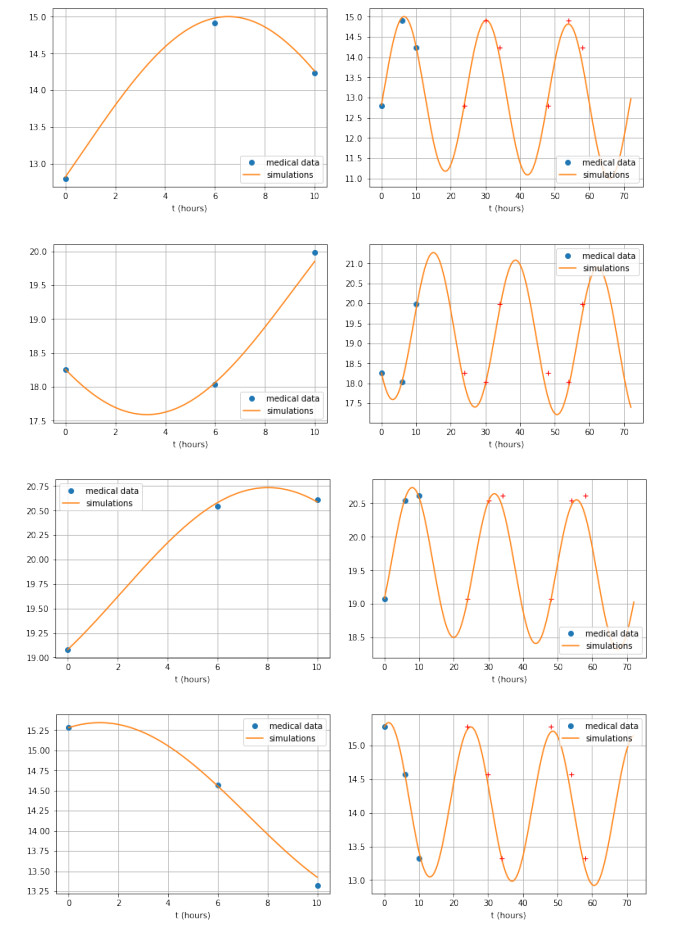
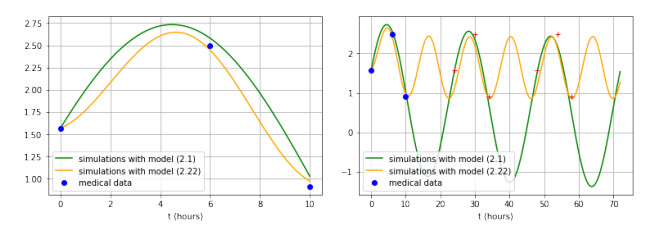
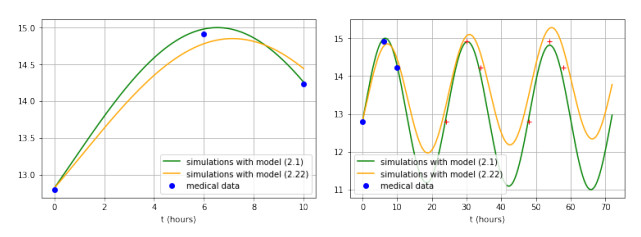
Figures(3)

Laura Hatchondo, Carole Guillevin, Mathieu Naudin, Laurence Cherfils, Alain Miranville, Rémy Guillevin. Mathematical modeling of brain metabolites variations in the circadian rhythm[J]. AIMS Mathematics, 2020, 5(1): 216-225. doi: 10.3934/math.2020013







 DownLoad:
DownLoad: