Citation: Albert Uchenna Ude, Che Husna Azhari. Lateral crashworthiness response of bombyx mori fibre/glass–fibre/epoxy hybrid composite cylindrical tubes-experimental[J]. AIMS Materials Science, 2019, 6(6): 1227-1239. doi: 10.3934/matersci.2019.6.1227

| [1] |
Mamalis AG, Manolakos DE, Ioannidis MB, et al. (2004) Crashworthy characteristics of axially statically compressed thin-walled square CFRP composite tubes: experimental. Compos Struct 63: 347-360. doi: 10.1016/S0263-8223(03)00183-1

|
| [2] |
Ude AU, Ariffin AK, Azhari CH (2013) Impact damage characteristics in reinforced woven natural silk/epoxy composite face-sheet and sandwich foam, coremat and honeycomb materials. Int J Impact Eng 58: 31-38. doi: 10.1016/j.ijimpeng.2013.03.003
|
| [3] |
Ude AU, Eshkoor RA, Azhari CH (2017) Crashworthy characteristics of axial quasi-statically compressed bombyx mori composite cylindrical tubes: experimental. Fiber Polym 18: 1594-1601. doi: 10.1007/s12221-017-1235-1
|
| [4] |
Supian ABM, Sapuan SM, Zuhri MYM, et al. (2018) Hybrid reinforced thermoset polymer composite in energy absorption tube application: A review. Def Technol 14: 291-305. doi: 10.1016/j.dt.2018.04.004
|
| [5] |
Eshkoor RA, Ude AU, Oshkovr SA, et al. (2014) Failure mechanism of woven natural silk/epoxy rectangular composite tubes under axial quasi-static crushing test using trigger mechanism. Int J Impact Eng 64: 53-61. doi: 10.1016/j.ijimpeng.2013.09.004
|
| [6] |
Eshkoor RA, Ude AU, Sulong AB, et al. (2015) Energy absorption and load carrying capability of woven natural silk epoxy-triggered composite tubes. Compos Part B-Eng 77: 10-18. doi: 10.1016/j.compositesb.2015.03.017
|
| [7] |
Eshkoor RA, Oshkovr SA, Sulong AB, et al. (2013) Effect of trigger configuration on the crashworthiness characteristics of natural silk epoxy composite tubes. Compos Part B-Eng 55: 5-10. doi: 10.1016/j.compositesb.2013.05.022
|
| [8] |
Eshkoor RA, Oshkovr SA, Sulong AB, et al. (2013) Comparative research on the crashworthiness characteristics of woven natural silk/epoxy composite tubes. Mater Des 47: 248-257. doi: 10.1016/j.matdes.2012.11.030
|
| [9] |
Cormier JR, LaPlante G (2018) Study of the effects of low-velocity impact on a composite bicycle down tube. Compos Struct 198: 144-155. doi: 10.1016/j.compstruct.2018.05.007
|
| [10] |
Kathiresan M, Manisekar K (2017) Low velocity axial collapse behavior of E-glass fiber/epoxy composite conical frusta. Compos Struct 166: 1-11. doi: 10.1016/j.compstruct.2017.01.041
|
| [11] |
Abdewi EF, Sulaiman S, Hamouda AMS, et al. (2008) Quasi-static axial and lateral crushing of radial corrugated composite tubes. Thin Wall Struct 46: 320-332. doi: 10.1016/j.tws.2007.07.018
|
| [12] |
Fan Z, Shen J, Lu G (2011) Investigation of lateral crushing of sandwich tubes. Procedia Eng 14: 442-449. doi: 10.1016/j.proeng.2011.07.055
|
| [13] |
Mahdi ES, El Kadi H (2008) Crushing behavior of laterally compressed composite elliptical tubes: experiments and predictions using artificial neural networks. Compos Struct 83: 399-412. doi: 10.1016/j.compstruct.2007.05.009
|
| [14] |
Abosbaia AS, Mahdi E, Hamouda AMS, et al. (2005) Energy absorption capability of laterally loaded segmented composite tubes. Compos Struct 70: 356-373. doi: 10.1016/j.compstruct.2004.08.039
|
| [15] |
Sebaey TA, Mahdi E (2016) Crashworthiness of pre-impacted glass/epoxy composite tubes. Int J Impact Eng 92: 18-25. doi: 10.1016/j.ijimpeng.2015.11.007
|
| [16] |
Moeinifard M, Liaghat G, Rahimi G, et al. (2016) Experimental investigation on the energy absorption and contact force of unstiffened and grid-stiffened composite cylindrical shells under lateral compression. Compos Struct 152: 626-36. doi: 10.1016/j.compstruct.2016.05.067
|
| [17] | Ali AM, Robillard D, Masmoudi R, et al. (2019) Experimental investigation of bond and tube thickness effect on the flexural behavior of concrete-filled FPR tube under lateral cyclic loading. J King Saud Univ Eng Sci 31: 32-41. |
| [18] |
Elahi SA, Rouzegar J, Niknejad A, et al. (2017) Theoretical study of absorbed energy by empty and foam-filled composite tubes under lateral compression. Thin Wall Struct 114: 1-10. doi: 10.1016/j.tws.2017.01.029
|
| [19] |
Liu Q, Xu X, Ma J, et al. (2017) Lateral crushing and bending responses of CFRP square tube filled with aluminum honeycomb. Compos Part B-Eng 118: 104-115. doi: 10.1016/j.compositesb.2017.03.021
|
| [20] |
Pol MH, Golshan NR (2019) Experimental investigation of parameters affected on behavior of composite tubes under quasi static and dynamic axial loading. Compos Part B-Eng 163: 471-486. doi: 10.1016/j.compositesb.2019.01.011
|
| [21] |
Cihan M, Sobey A, Blake JIR (2019) Mechanical and dynamic performance of woven flax/E-glass hybrid composites. Compos Sci Technol 172: 36-42. doi: 10.1016/j.compscitech.2018.12.030
|
| [22] |
Mamalis AG, Manolakos DE, Ioannidis MB, et al. (2005) On the response of thin-walled CFRP composite tubular components subjected to static and dynamic axial compressive loading: experimental. Compos Struct 69: 407-420. doi: 10.1016/j.compstruct.2004.07.021
|

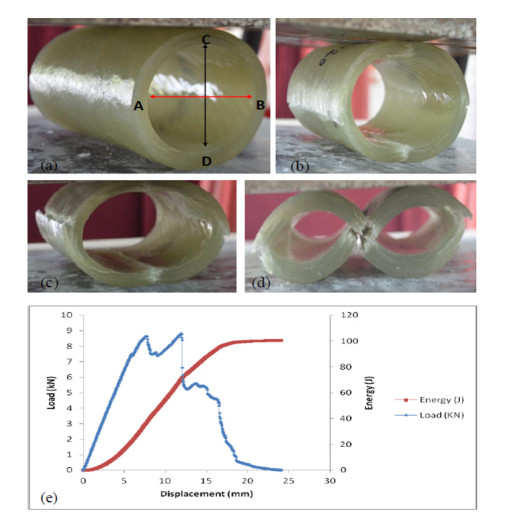
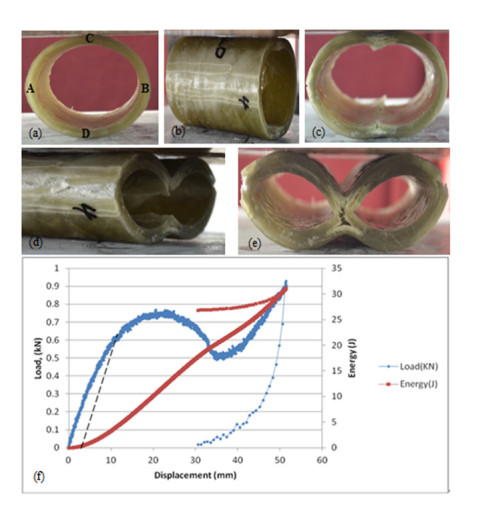
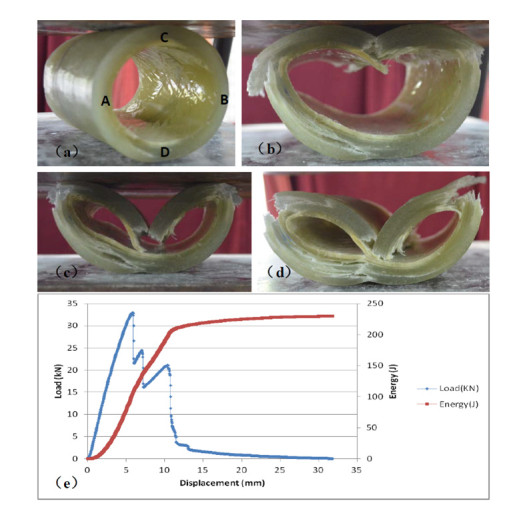
Figures(11)

Albert Uchenna Ude, Che Husna Azhari. Lateral crashworthiness response of bombyx mori fibre/glass–fibre/epoxy hybrid composite cylindrical tubes-experimental[J]. AIMS Materials Science, 2019, 6(6): 1227-1239. doi: 10.3934/matersci.2019.6.1227







 DownLoad:
DownLoad: