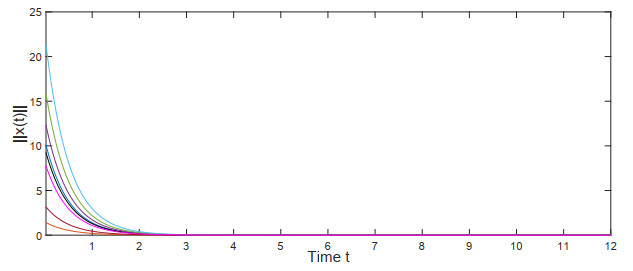
This manuscript investigated the stability of Hopfield neural networks with time-varying transmission delays and leakage delays, and specially discussed the impact of the time-varying delays' dynamic characteristics on network stability. First, to characterize the dynamic features of time-varying delays transitioning continuously between short and large delays, two key parameters were introduced: a critical threshold for distinguishing whether the time-varying delays are short or large delays, and the ratio of the measure of the union of time periods, in which the time-varying delays appear as short delays, to the measure of the whole time interval. Then, by utilizing the Lyapunov stability method, some sufficient conditions for the global exponential stability of the neural networks were derived. It is pointed out that when the measure of time periods in which the time-varying delays appear as short delays is large enough, the above two parameters related to the time-varying delays' dynamic characteristics will have an important impact on the system stability, and the upper bound that the time-varying delays can achieve in the whole time interval will no longer be the dominant factor influencing stability. Additionally, the relationship between the leakage delays and the stabilization ability of the negative feedback terms was explored. Two admissible upper bounds were presented, below which the leakage delays do not completely undermine the capacity of negative feedback terms to stabilize the neural networks. Finally, some simulation experiments were conducted to validate our theoretical findings.
Citation: Xudong Hai, Chengxu Chen, Qingyun Wang, Xiaowei Ding, Zhaoying Ye, Yongguang Yu. Effect of time-varying delays' dynamic characteristics on the stability of Hopfield neural networks[J]. Electronic Research Archive, 2025, 33(3): 1207-1230. doi: 10.3934/era.2025054
This manuscript investigated the stability of Hopfield neural networks with time-varying transmission delays and leakage delays, and specially discussed the impact of the time-varying delays' dynamic characteristics on network stability. First, to characterize the dynamic features of time-varying delays transitioning continuously between short and large delays, two key parameters were introduced: a critical threshold for distinguishing whether the time-varying delays are short or large delays, and the ratio of the measure of the union of time periods, in which the time-varying delays appear as short delays, to the measure of the whole time interval. Then, by utilizing the Lyapunov stability method, some sufficient conditions for the global exponential stability of the neural networks were derived. It is pointed out that when the measure of time periods in which the time-varying delays appear as short delays is large enough, the above two parameters related to the time-varying delays' dynamic characteristics will have an important impact on the system stability, and the upper bound that the time-varying delays can achieve in the whole time interval will no longer be the dominant factor influencing stability. Additionally, the relationship between the leakage delays and the stabilization ability of the negative feedback terms was explored. Two admissible upper bounds were presented, below which the leakage delays do not completely undermine the capacity of negative feedback terms to stabilize the neural networks. Finally, some simulation experiments were conducted to validate our theoretical findings.

| [1] |
J. J. Hopfield, Neural networks and physical systems with emergent collective computational abilities, Proc. Natl. Acad. Sci. U.S.A., 79 (1982), 2554–2558. https://doi.org/10.1073/pnas.79.8.2554 doi: 10.1073/pnas.79.8.2554

|
| [2] |
S. K. Duan, X. F. Hu, Z. K. Dong, L. D. Wang, P. Mazumder, Memristor-based cellular nonlinear/neural network: Design, analysis, and applications, IEEE Trans. Neural Networks Learn. Syst., 26 (2015), 1202–1213. https://doi.org/10.1109/TNNLS.2014.2334701 doi: 10.1109/TNNLS.2014.2334701
|
| [3] |
Z. G. Zeng, J. Wang, Associative memories based on continuous-time cellular neural networks designed using space-invariant cloning templates, Neural Networks, 22 (2009), 651–657. https://doi.org/10.1016/j.neunet.2009.06.031 doi: 10.1016/j.neunet.2009.06.031
|
| [4] |
A. Ascoli, V. Lanza, F. Corinto, R. Tetzlaff, Synchronization conditions in simple memristor neural networks, J. Franklin Inst., 352 (2015), 3196–3220. https://doi.org/10.1016/j.jfranklin.2015.06.003 doi: 10.1016/j.jfranklin.2015.06.003
|
| [5] |
E. Aktas, O. Faydasicok, S. Arik, Robust stability of dynamical neural networks with multiple time delays: A review and new results, Artif. Intell. Rev., 56 (2023), 1647–1684. https://doi.org/10.1007/s10462-023-10552-x doi: 10.1007/s10462-023-10552-x
|
| [6] |
X. Y. Zhang, X. D. Li, J. D. Cao, F. Miaadi, Design of memory controllers for finite-time stabilization of delayed neural networks with uncertainty, Artif. Intell. Rev., 355 (2018), 5394–5413. https://doi.org/10.1016/j.jfranklin.2018.05.037 doi: 10.1016/j.jfranklin.2018.05.037
|
| [7] |
Z. Ma, G. Sun, D. Liu, X. Xing, Dissipativity analysis for discrete-time fuzzy neural networks with leakage and time-varying delays, Neurocomputing, 175 (2016), 579–584. https://doi.org/10.1016/j.neucom.2015.10.098 doi: 10.1016/j.neucom.2015.10.098
|
| [8] |
W. T. Hua, Y. T. Wang, C. Y. Liu, New method for global exponential synchronization of multi-link memristive neural networks with three kinds of time-varying delays, Appl. Math. Comput., 471 (2024), 128593. https://doi.org/10.1016/j.amc.2024.128593 doi: 10.1016/j.amc.2024.128593
|
| [9] |
Q. K. Song, H. Q. Shu, Z. J. Zhao, Y. R. Liu, F. E. Alsaadi, Lagrange stability analysis for complex-valued neural networks with leakage delay and mixed time-varying delays, Neurocomputing, 244 (2017), 33–41. https://doi.org/10.1016/j.neucom.2017.03.015 doi: 10.1016/j.neucom.2017.03.015
|
| [10] | K. Gopalsamy, Leakage delays in BAM, J. Math. Anal. Appl., 325 (2007), 1117–1132. https://doi.org/10.1016/j.jmaa.2006.02.039 |
| [11] |
X. M. Zhang, Q. L. Han, State estimation for static neural networks with time-varying delays based on an improved reciprocally convex inequality, IEEE Trans. Neural Networks Learn. Syst., 29 (2018), 1376–1381. https://doi.org/10.1109/TNNLS.2017.2661862 doi: 10.1109/TNNLS.2017.2661862
|
| [12] |
M. Sader, A. Abdurahman, H. J. Jiang, General decay lag synchronization for competitive neural networks with constant delays, Neural Process. Lett., 50 (2019), 445–457. https://doi.org/10.1007/s11063-019-09984-w doi: 10.1007/s11063-019-09984-w
|
| [13] |
Y. Wang, C. D. Zheng, E. Feng, Stability analysis of mixed recurrent neural networks with time delay in the leakage term under impulsive perturbations, Neurocomputing, 119 (2013), 454–461. https://doi.org/10.1016/j.neucom.2013.03.012 doi: 10.1016/j.neucom.2013.03.012
|
| [14] |
Y. Chen, Z. Fu, Y. Liu, F. E. Alsaadi, Further results on passivity analysis of delayed neural networks with leakage delay, Neurocomputing, 224 (2017), 135–141. https://doi.org/10.1016/j.neucom.2016.10.045 doi: 10.1016/j.neucom.2016.10.045
|
| [15] |
X. Y. Zhang, D. G. Wang, B. Yang, K. R. Ota, M. X. Dong, H. X. Li, Exponentially synchronous results for delayed neural networks with leakage delay via switched delay idea and AED-ADT method, IEEE Trans. Neural Networks Learn. Syst., 35 (2024), 3713–3724. https://doi.org/10.1109/TNNLS.2022.3196402 doi: 10.1109/TNNLS.2022.3196402
|
| [16] |
C. R. Wang, F. Long, K. Y. Xie, H. T. Wang, C. K. Zhang, Y. He, A degree-dependent polynomial-based reciprocally convex matrix inequality and its application to stability analysis of delayed neural networks, IEEE Tran. Sybernetics, 54 (2024), 4164–4176. https://doi.org/10.1109/TCYB.2024.3365709 doi: 10.1109/TCYB.2024.3365709
|
| [17] |
H. G. Zhang, D. W. Gong, B. Chen, Z. W. Liu, Synchronization for coupled neural networks with interval delay: A novel augmented Lyapunov-Krasovskii functional method, IEEE Trans. Neural Networks Learn. Syst., 24 (2013), 58–70. https://doi.org/10.1109/TNNLS.2012.2225444 doi: 10.1109/TNNLS.2012.2225444
|
| [18] |
B. Zhang, S. Xu, Y. Zou, Improved delay-dependent exponential stability criteria for discrete-time recurrent neural networks with time-varying delays, Neurocomputing, 72 (2008), 321–330. https://doi.org/10.1016/j.neucom.2008.01.006 doi: 10.1016/j.neucom.2008.01.006
|
| [19] |
P. Park, J. Ko, C. Jeong, Reciprocally convex approach to stability of systems with time-varying delays, Automatica, 47 (2011), 235–238. https://doi.org/10.1016/j.automatica.2010.10.014 doi: 10.1016/j.automatica.2010.10.014
|
| [20] |
Q. X. Zhu, J. D. Cao, Exponential stability of stochastic neural networks with both markovian jump parameters and mixed time delays, IEEE Tran. Syst. Man Cybern. Part B, 41 (2011), 341–353. https://doi.org/10.1109/TSMCB.2010.2053354 doi: 10.1109/TSMCB.2010.2053354
|
| [21] |
H. B. Zeng, J. H. Park, C. F. Zhang, W. Wang, Stability and dissipativity analysis of static neural networks with interval time-varying delay, J. Franklin Inst., 352 (2015), 1284–1295. https://doi.org/10.1016/j.jfranklin.2014.12.023 doi: 10.1016/j.jfranklin.2014.12.023
|
| [22] |
H. C. Lin, H. B. Zeng, X. M. Zhang, W. Wang, Stability analysis for delayed neural networks via a generalized reciprocally convex inequality, IEEE Trans. Neural Networks Learn. Syst., 34 (2023), 7491–7499. https://doi.org/10.1109/TNNLS.2022.3144032 doi: 10.1109/TNNLS.2022.3144032
|
| [23] |
X. Wei, Z. Zhang, M. Liu, Z. Wang, J. Chen, Anti-synchronization for complex-valued neural networks with leakage delay and time-varying delays, Neurocomputing, 412 (2020), 312–319. https://doi.org/10.1016/j.neucom.2020.06.080 doi: 10.1016/j.neucom.2020.06.080
|
| [24] |
H. B. Zeng, Z. J. Zhu, W. Wang, X. M. Zhang, Relaxed stability criteria of delayed neural networks using delay-parameters-dependent slack matrices, Neural Networks, 180 (2024), 106676. https://doi.org/10.1016/j.neunet.2024.106676 doi: 10.1016/j.neunet.2024.106676
|
| [25] |
X. M. Zhang, Q. L. Han, Z. G. Zeng, Hierarchical type stability criteria for delayed neural networks via canonical bessel-legendre inequalities, IEEE Trans. Cybern., 48 (2018), 1660–1671. https://doi.org/10.1109/TCYB.2017.2776283 doi: 10.1109/TCYB.2017.2776283
|
| [26] |
Q. X. Zhu, J. D. Cao, Robust exponential stability of markovian jump impulsive stochastic Cohen-Grossberg neural networks with mixed time delays, IEEE Trans. Neural Networks, 21 (2010), 1314–1325. https://doi.org/10.1109/TNN.2010.2054108 doi: 10.1109/TNN.2010.2054108
|
| [27] |
X. Zhang, S. L. Yuan, Y. T. Wang, X. A. Yang, A novel direct method to H$_{\infty}$ synchronization of switching inertial neural networks with mixed time-varying delays, Neurocomputing, 598 (2024), 128039. https://doi.org/10.1016/j.neucom.2024.128039 doi: 10.1016/j.neucom.2024.128039
|
| [28] |
Y. Chen, Y. Q. Li, G. Chen, New results on stability analysis for a class of generalized delayed neural networks, Appl. Math. Comput., 469 (2024), 128529. https://doi.org/10.1016/j.amc.2024.128529 doi: 10.1016/j.amc.2024.128529
|
| [29] |
X. L. Qin, C. Wang, L. X. Li, H. P. Peng, Y. X. Yang, L. Ye, Finite-time modified projective synchronization of memristor-based neural network with multi-links and leakage delay, Chaos Solitons Fractals, 116 (2018), 302–315. https://doi.org/10.1016/j.chaos.2018.09.040 doi: 10.1016/j.chaos.2018.09.040
|
| [30] |
J. W. Cao, S. M. Zhong, Y. Y. Hu, Global stability analysis for a class of neural networks with varying delays and control input, Appl. Math. Comput., 189 (2007), 1480–1490. https://doi.org/10.1016/j.amc.2006.12.048 doi: 10.1016/j.amc.2006.12.048
|
| [31] |
T. Ensari, S. Arik, Global stability analysis of neural networks with multiple time varying delays, IEEE Trans. Autom. Control, 50 (2005), 1781–1785. https://doi.org/10.1109/TAC.2005.858634 doi: 10.1109/TAC.2005.858634
|

Figures(2)
Xudong Hai, Chengxu Chen, Qingyun Wang, Xiaowei Ding, Zhaoying Ye, Yongguang Yu. Effect of time-varying delays' dynamic characteristics on the stability of Hopfield neural networks[J]. Electronic Research Archive, 2025, 33(3): 1207-1230. doi: 10.3934/era.2025054







 DownLoad:
DownLoad: