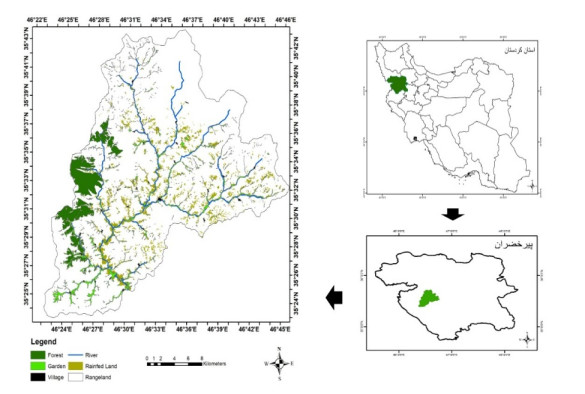
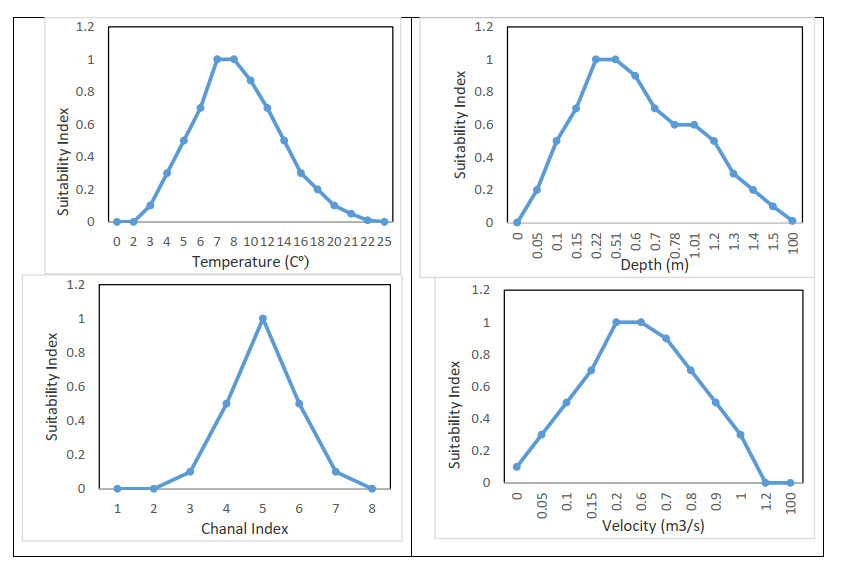
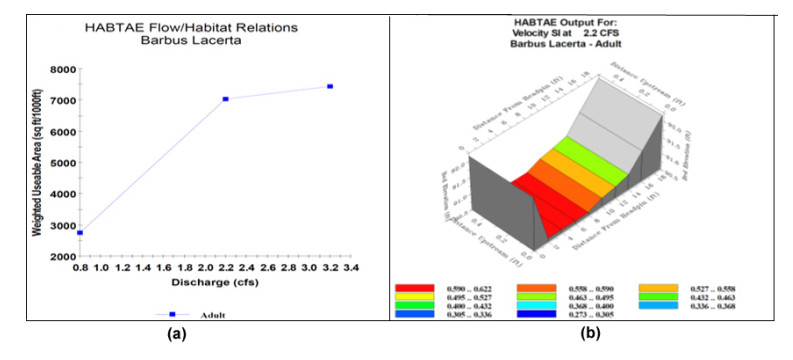
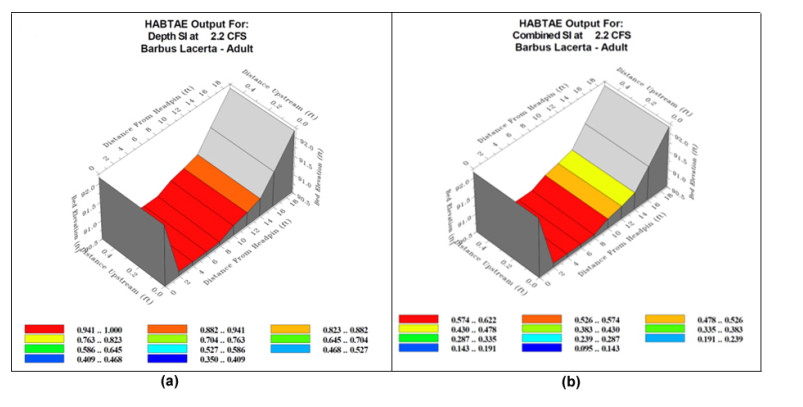
Proper water resource management is essential for maintaining a sustainable supply chain and meeting water demand. The urgent need to preserve river ecosystems by sustaining environmental flow (EF) in the realm of environmental management has been highlighted by the drastic changes to river ecosystems and upstream flow dynamics brought about by careless river exploitation in the last few decades. To optimize EF in river basin management, we present an integrated modeling approach. We focused on the Pir Khezran River basin. Our objective was to estimate EF and generalize the findings to adjacent rivers using modeling techniques, thus providing valuable insights for environmental management applications. The assessment and optimization of EF under uncertain conditions was achieved by combining physical habitat simulation (PHABSIM) modeling with advanced techniques like Adaptive Neuro-Fuzzy Inference Systems (ANFIS) and Multilayer Perceptron (MLP) neural networks. This integrated modeling approach contributes to sustainable solutions for river basin management and environmental conservation by effectively optimizing EF, as demonstrated by the results. This research, therefore, makes valuable contributions to environmental management in various areas such as ecological preservation, modeling and optimizing environmental systems, and policy considerations.
Citation: Seiran Haghgoo, Jamil Amanollahi, Barzan Bahrami Kamangar, Shahryar Sorooshian. Decision models enhancing environmental flow sustainability: A strategic approach to water resource management[J]. AIMS Environmental Science, 2024, 11(6): 900-917. doi: 10.3934/environsci.2024045
Proper water resource management is essential for maintaining a sustainable supply chain and meeting water demand. The urgent need to preserve river ecosystems by sustaining environmental flow (EF) in the realm of environmental management has been highlighted by the drastic changes to river ecosystems and upstream flow dynamics brought about by careless river exploitation in the last few decades. To optimize EF in river basin management, we present an integrated modeling approach. We focused on the Pir Khezran River basin. Our objective was to estimate EF and generalize the findings to adjacent rivers using modeling techniques, thus providing valuable insights for environmental management applications. The assessment and optimization of EF under uncertain conditions was achieved by combining physical habitat simulation (PHABSIM) modeling with advanced techniques like Adaptive Neuro-Fuzzy Inference Systems (ANFIS) and Multilayer Perceptron (MLP) neural networks. This integrated modeling approach contributes to sustainable solutions for river basin management and environmental conservation by effectively optimizing EF, as demonstrated by the results. This research, therefore, makes valuable contributions to environmental management in various areas such as ecological preservation, modeling and optimizing environmental systems, and policy considerations.

| [1] |
Pandey CL (2021) Managing urban water security: challenges and prospects in Nepal. Environ Dev Sustain 23: 241–257. https://doi.org/10.1007/s10668-019-00577-0 doi: 10.1007/s10668-019-00577-0

|
| [2] | IUCN (World Conservation Union) (2000). Vision for Water and Nature. A World Strategy for conservation and sustainable management of water resources in the 21st Century. IUCN, Gland, Switzerland and Cambridge, UK, p 52. |
| [3] |
Perez-Blanco CD, Gil-Garcia L, Saiz-Santiago (2021) An actionable hydroeconomic descision support system for the assessment of water reallocations in irrigated agriculture. Astudy of minimum environmental flows in the Douro River Basin, Spain. J Environ Managet 298: 113432. https://doi.org/10.1016/j.jenvman.2021.113432 doi: 10.1016/j.jenvman.2021.113432
|
| [4] |
Senent-Aparicio J, George C, Srinivasan R (2021) Introducing a new post-processing tool for the SWAT+model to evaluate environmental flows. Environ Modell Softw 136: 104944. https://doi.org/10.1016/j.envsoft.2020.104944 doi: 10.1016/j.envsoft.2020.104944
|
| [5] |
Zolfagharpour F, Saghafian B, Delavar M (2021) Adapting reservoir operation rules to hydrological drought state and environmental flow requirements. J Hydrol 660: 126581. https://doi.org/10.1016/j.jhydrol.2021.126581 doi: 10.1016/j.jhydrol.2021.126581
|
| [6] |
Sedighkia M, Datta B (2023) Analyzing environmental flow supply in the semi-arid area through integrating drought analysis and optimal operation of reservoir. J Arid Land 15: 1439–1454. https://doi.org/10.1007/s40333-023-0035-2 doi: 10.1007/s40333-023-0035-2
|
| [7] |
Aazami J, Motevalli A, Savabieasfahani M (2022) Correction to: Evaluation of three environmental flow techniques in Shoor wetland of Golpayegan, Iran. Int J Environ Sci Technol 19: 7899. https://doi.org/10.1007/s13762-022-04289-3 doi: 10.1007/s13762-022-04289-3
|
| [8] | Wang H, Cong P, Zhu Z, et al. (2022) Analysis of environmental dispersion in wetland flows with floating vegetation islands. J Hydrol 606: 127359. https://doi.org/10.1016/j.jhydrol.2021.127359 |
| [9] |
Arthington ÁH, Naiman RJ, Mcclain ME, et al. (2010). Preserving the biodiversity and ecological services of rivers: new challenges and research opportunities. Freshwater Biol 55: 1–16. https://doi.org/10.1111/j.1365-2427.2009.02340.x doi: 10.1111/j.1365-2427.2009.02340.x
|
| [10] |
Chilton D, Hamilton DP, Nagelkerken I, et al. (2021). Environmental flow requirements of estuaries: providing resilience to current and future climate and direct anthropogenic changes. Front Environ Sci 9: 764218. https://doi.org/10.3389/fenvs.2021.764218 doi: 10.3389/fenvs.2021.764218
|
| [11] |
Espinoza T, Burke CL, Carpenter-Bundhoo L, et al. (2021) Quantifying movement of multiple threatened species to inform adaptive management of environmental flows. J Environ Managet 295: 113067. https://doi.org/10.1016/j.jenvman.2021.113067 doi: 10.1016/j.jenvman.2021.113067
|
| [12] |
Zhang Y, Yu H, Yu H, et al. (2020) Optimization of environmental variables in habitat suitability modeling for mantis shrimp Oratosquilla oratoria in the Haizhou Bay and adjacent waters. Acta Oceanol Sin 39: 36–47. https://doi.org/10.1007/s13131-020-1546-8 doi: 10.1007/s13131-020-1546-8
|
| [13] |
Vivian LM, Godfree RC, Colloff MJ, et al. (2014) Wetland plant growth under contrasting water regimes associated with river regulation and drought: implications for environmental water management. Plant Ecol 215: 997–1011. https://doi.org/10.1007/s11258-014-0357-4 doi: 10.1007/s11258-014-0357-4
|
| [14] |
Gain AK, Apel H, Renaud FG, et al. (2013) Thresholds of hydrologic flow regime of a river and investigation of climate change impact—the case of the Lower Brahmaputra river Basin. Climatic Change 120: 463–475. https://doi.org/10.1007/s10584-013-0800-x doi: 10.1007/s10584-013-0800-x
|
| [15] |
Belmar O, Velasco J, Martinez-Capel F (2011) Hydrological Classification of Natural Flow Regimes to Support Environmental Flow Assessments in Intensively Regulated Mediterranean Rivers, Segura River Basin (Spain). Environ Manage 47: 992–1004. https://doi.org/10.1007/s00267-011-9661-0 doi: 10.1007/s00267-011-9661-0
|
| [16] |
Dumitriu D (2020) Sediment flux during flood events along the Trotuș River channel: hydrogeomorphological approach. J Soils Sediments 20: 4083–4102. https://doi.org/10.1007/s11368-020-02763-4 doi: 10.1007/s11368-020-02763-4
|
| [17] |
Bouska K (2020) Regime change in a large-floodplain river ecosystem: patterns in body-size and functional biomass indicate a shift in fish communities. Biol Invasions 22: 3371–3389. https://doi.org/10.1007/s10530-020-02330-5 doi: 10.1007/s10530-020-02330-5
|
| [18] |
Poff NL, Allan JD, Bain MB, et al. (1997). The natural flow regime. Bio Science 47: 769–784. https://doi.org/10.2307/1313099 doi: 10.2307/1313099
|
| [19] | Nasiri Khiavi A, Mostafazadeh R, Ghanbari Talouki F (2024) Using game theory algorithm to identify critical watersheds based on environmental flow components and hydrological indicators. Environ Dev Sustain https://doi-org.access.semantak.com/10.1007/s10668-023-04390-8 |
| [20] |
Sedighkia M, Abdoli A (2023) Design of optimal environmental flow regime at downstream of multireservoir systems by a coupled SWAT-reservoir operation optimization method. Environ Dev Sustain 25: 834–854. https://doi.org/10.1007/s10668-021-02081-w doi: 10.1007/s10668-021-02081-w
|
| [21] |
Yu L, Wu X, Wu S, et al. (2021) Multi-objective optimal operation of cascade hydropower plants considering ecological flow under different ecological conditions. J Hydrol 601: 126599. https://doi.org/10.1016/j.jhydrol.2021.126599 doi: 10.1016/j.jhydrol.2021.126599
|
| [22] |
Mlynski D, Operacz A, Walega A (2020) Sensitivity of methods for calculating environmental flows based on hydrological characteristics of watercourses regarding te hydropower potential of rivers. J Clean Pro 250: 119527. https://doi.org/10.1016/j.jclepro.2019.119527 doi: 10.1016/j.jclepro.2019.119527
|
| [23] |
Bejarano MD, Sord-Ward A, Gabriel-Martin I, et al. (2019) Tradeoff between economic and environmental costs and benefits of hydropower production at run-of-river-diversion schemes under different environmental flows scenarios. J Hydrol 572: 790–804. https://doi.org/10.1016/j.jhydrol.2019.03.048 doi: 10.1016/j.jhydrol.2019.03.048
|
| [24] | Bovee KD (1982) Instream flow methodology. US Fish and Wildlife Service. FWS/OBS, 82, 26 |
| [25] |
Stucchi L. Bocchiola D (2023) Environmental Flow Assessment using multiple criteria: A case study in the Kumbih river, West Sumatra (Indonesia). Sci Total Environ 901: 166516. https://doi.org/10.1016/j.scitotenv.2023.166516 doi: 10.1016/j.scitotenv.2023.166516
|
| [26] |
Mwamila TB, Kimwaga RJ, Mtalo FW (2008) Eco-hydrology of the Pangani River downstream of Nyumba ya Mungu reservoir, Tanzania. Phys Chem Earth 33: 695–700. https://doi.org/10.1016/j.pce.2008.06.054 doi: 10.1016/j.pce.2008.06.054
|
| [27] |
Nagaya T, Shiraishi Y, Onitsuka K, et al (2008) Evaluation of suitable hydraulic conditions for spawning of ayu with horizontal 2D numerical simulation and PHABSIM. Ecol Modell 215: 133–143. https://doi.org/10.1016/j.ecolmodel.2008.02.043 doi: 10.1016/j.ecolmodel.2008.02.043
|
| [28] |
Nikghalb S, Shokoohi A, Singh VP et al (2016) Ecological Regime versus Minimum Environmental Flow:Comparison of Results for a River in a Semi Mediterranean Region. Water Resour Manage 30: 4969–4984. https://doi.org/10.1007/s11269-016-1488-2 doi: 10.1007/s11269-016-1488-2
|
| [29] |
Spence R, Hickley P (2000) The use of PHABSIM in the management of water resources and fisheries in England and Wales. Ecol Eng 16: 153–158. https://doi.org/10.1016/S0925-8574(00)00099-9 doi: 10.1016/S0925-8574(00)00099-9
|
| [30] |
Miao Y, Li J, Feng P, et al. (2020) Effects of land use changes on the ecological operation of the Panjiakou-Daheiting Reservoir system, China Ecol Eng 152: 10585. https://doi.org/10.1016/j.ecoleng.2020.105851 doi: 10.1016/j.ecoleng.2020.105851
|
| [31] |
Dana K, Jana C, Tomáš V (2017) Analysis of environmental flow requirements for macroinvertebrates in a creek affected by urban drainage (Prague metropolitan area, Czech Republic). Urban Ecosyst 20: 785–797. https://doi.org/10.1007/s11252-017-0649-2 doi: 10.1007/s11252-017-0649-2
|
| [32] |
Weng X, Jiang C, Yuan M, et al. (2021) An ecologically dispatch strategy using environmental flows for a cascade multi-sluice system: A case study of the Yongjiang River Basin, China Ecol Indic 121: 107053. https://doi.org/10.1016/j.ecolind.2020.107053 doi: 10.1016/j.ecolind.2020.107053
|
| [33] |
Knack IM, Huang F, Shen HT (2020) Modeling fish habitat condition in ice affected rivers. Cold Reg Sci Technol 176: 103086. https://doi.org/10.1016/j.coldregions.2020.103086 doi: 10.1016/j.coldregions.2020.103086
|
| [34] |
Peng L, Sun L (2016) Minimum instream flow requirement for the water-reduction section of diversion-type hydropower station: a case study of the Zagunao River, China. Environ Earth Sci 75: 1210. https://doi.org/10.1007/s12665-016-6019-1 doi: 10.1007/s12665-016-6019-1
|
| [35] |
Gholami V, Khalili A, Sahour H, et al. (2020) Assessment of environmental water requirement for rivers of the Miankaleh wetland drainage basin. Appl Water Sci 10: 233. https://doi.org/10.1007/s13201-020-01319-8 doi: 10.1007/s13201-020-01319-8
|
| [36] |
Im D, Choi SU, Choi B (2018) Physical habitat simulation for a fish community using the ANFIS method. Ecol Inform 43: 73-83. https://doi.org/10.1016/j.ecoinf.2017.09.001 doi: 10.1016/j.ecoinf.2017.09.001
|
| [37] |
Zamanzad-Ghavidel S, Fazeli S, Mozaffari S, et al. (2023) Estimating of aqueduct water withdrawal via a wavelet-hybrid soft-computing approach under uniform and non-uniform climatic conditions. Environ Dev Sustain 25: 5283–5314. https://doi.org/10.1007/s10668-022-02265-y doi: 10.1007/s10668-022-02265-y
|
| [38] | Emadi A, Sobhani R, Ahmadi H, et al. Multivariate modeling of agricultural river water abstraction via novel integrated-wavelet methods in various climatic conditions. Environ Dev Sustain 24: 4845–4871. https://doi.org/10.1007/s10668-021-01637-0 |
| [39] | Amanollahi J, Ausati Sh (2020a) PM2.5 concentration forecasting using ANFIS, EEMD-GRNN, MLP, and MLR models: A case study of Tehran, Iran. Air Qual Atmos Health 13: 161–17. https://doi.org/10.1007/s11869-019-00779-5 |
| [40] |
Ghasemi A, Amanollahi J (2019) Integration of ANFIS model and forward selection method for air quality forecasting. Air Qual Atmos Health 12: 59–72. https://doi.org/10.1007/s11869-018-0630-0 doi: 10.1007/s11869-018-0630-0
|
| [41] |
Kaboodvandpour S, Amanollahi J, Qhavami S, et al. (2015) Assessing the accuracy of multiple regressions, ANFIS, and ANN models in predicting dust storm occurrences in Sanandaj. Iran Nat Hazards 78: 879–893. https://doi.org/10.1007/s11069-015-1748-0 doi: 10.1007/s11069-015-1748-0
|
| [42] |
Bice CM, Gehrig SL, Zampatti BP, et al. (2014) Flow-induced alterations to fish assemblages, habitat and fish–habitat associations in a regulated lowland river. Hydrobiologia 722: 205–222. https://doi.org/10.1007/s10750-013-1701-8 doi: 10.1007/s10750-013-1701-8
|
| [43] |
Kim HJ, Kim JH, Ji U, et al. (2020) Effect of Probability Distribution-Based Physical Habitat Suitability Index on Environmental-Flow Estimation. KSCE. J Civ Eng 24: 2393–2402. https://doi.org/10.1007/s12205-020-1923-z doi: 10.1007/s12205-020-1923-z
|
| [44] | EPA, 2015. Fish Field Sampling. SESDPROC-512-R4. |
| [45] | Standard Methods for the Examination of Water and Wastewater, 1999. 20th Edition, Part 10600 B, Electrofishing, P 2206. |
| [46] |
Van der Lee GEM, Van der Molen DT, Van den Boogaard HFP, et al. (2006) Uncertainty analysis of a spatial habitat suitability model and implications for ecological management of water bodies. Landscape Ecol 21: 1019–1032. https://doi.org/10.1007/s10980-006-6587-7 doi: 10.1007/s10980-006-6587-7
|
| [47] |
Sekine M, Wang J, Yamamoto K, et al. (2020) Fish habitat evaluation based on width-to-depth ratio and eco-environmental diversity index in small rivers. Environ Sci Pollut Res 27: 34781–34795. https://doi.org/10.1007/s11356-020-08691-7 doi: 10.1007/s11356-020-08691-7
|
| [48] |
Wen X, Lv Y, Liu Z, et al. (2021) Operation chart optimization of multi-hydropower system incorporating the long- and short-term fish habitat requirements. J Clen Pro 121: 107053. https://doi.org/10.1016/j.jclepro.2020.125292 doi: 10.1016/j.jclepro.2020.125292
|
| [49] |
Jung SH, Choi S-Uk (2015) Prediction of composite suitability index for physical habitatsimulations using the ANFIS method. Appl Soft Comput 34: 502–512. https://doi.org/10.1016/j.asoc.2015.05.028 doi: 10.1016/j.asoc.2015.05.028
|
| [50] |
Fraternali P, Castelletti A, Soncini-Sessa R, et al. (2012) Putting humans in the loop: social computing for Water Resources Management. Environ Modell Softw 37: 68–77. https://doi.org/10.1016/j.envsoft.2012.03.002 doi: 10.1016/j.envsoft.2012.03.002
|
| [51] |
Rinderknecht SL, Borsuk ME, Reichert P (2012) Bridging uncertain and ambiguous knowledge with imprecise probabilities. Environ Modell Softw 36: 122–130. https://doi.org/10.1016/j.envsoft.2011.07.022 doi: 10.1016/j.envsoft.2011.07.022
|
| [52] |
Fukuda S (2009) Consideration of fuzziness: Is it necessary in modelling fish habitat preference of Japanese medaka (Oryzias latipes)? Ecol Model 220: 2877–2884. https://doi.org/10.1016/j.ecolmodel.2008.12.025 doi: 10.1016/j.ecolmodel.2008.12.025
|
| [53] |
Fukuda S, Hiramatsu B (2008) Prediction ability and sensitivity of artificial intelligence-based habitat preference models for predicting spatial distribution of Japanese medaka (Oryzias latipes). Ecol Model 215: 301–313. https://doi.org/10.1016/j.ecolmodel.2008.03.022 doi: 10.1016/j.ecolmodel.2008.03.022
|
| [54] |
Dawson, CW, Wilby R L (2001) Hydrological modelling using artificial neural networks. Progr Phys Geogr 25: 80–108. https://doi.org/10.1177/030913330102500104 doi: 10.1177/030913330102500104
|
| [55] |
Sykes AO (1993) An Introduction to Regression Analysis. Am Stat 61: 101. https://doi.org/10.1198/tas.2007.s74 doi: 10.1198/tas.2007.s74
|
| [56] | Kayes I, Shahriar SA, Hasan K, et al. (2019) The relationships between meteorological parameters and air pollutants in an urban environment. Global J Environ Sci Manag 5: 265–278. |
| [57] |
Ceylan Ζ, Bulkan S (2018) Forecasting PM10 levels using ANN and MLR: A case study for Sakarya City. Global Nest J 20: 281–290. https://doi.org/10.30955/gnj.002522 doi: 10.30955/gnj.002522
|
| [58] |
Karatzas K, Katsifarakis N, Orlowski C, et al. (2018) Revisiting urban air quality forecasting: a regression approach. Vietnam J Comput Sci 5: 177–184. https://doi.org/10.1007/s40595-018-0113-0 doi: 10.1007/s40595-018-0113-0
|
| [59] |
Mishra D, Goyal P (2015) Development of artificial intelligence based NO2 forecasting models at Taj Mahal, Agra. Atmos Pollut Res 6: 99–106. https://doi.org/10.5094/APR.2015.012 doi: 10.5094/APR.2015.012
|
| [60] |
Mishra D, Goyal P (2016) Neuro-Fuzzy approach to forecasting ozone episodes over the urban area of Delhi, India. Environ Technol Inno 5: 83–94. https://doi.org/10.1016/j.eti.2016.01.001 doi: 10.1016/j.eti.2016.01.001
|
| [61] |
Rahimi A (2017) Short-term prediction of NO 2 and NO x concentrations using multilayer perceptron neural network: a case study of Tabriz, Iran. Ecol Process 6: 4. https://doi.org/10.1186/s13717-016-0069-x doi: 10.1186/s13717-016-0069-x
|
| [62] |
Abba S, Hadi SJ, Abdullahi J (2017) River water modelling prediction using multi linear regression, artificial neural network, and adaptive neuro-fuzzy inference system techniques. Procedia Comput Sci 120: 75–82. https://doi.org/10.1016/j.procs.2017.11.212 doi: 10.1016/j.procs.2017.11.212
|
| [63] |
Abba SI, Elkirn G (2017) Effluent prediction of chemical oxygen demand from the astewater treatment plant using artificial neural network application. Procedia Comput Sci 120: 156–163. https://doi.org/10.1016/j.procs.2017.11.223 doi: 10.1016/j.procs.2017.11.223
|
| [64] |
Mohammadi S, Naseri F, Abri R (2019) Simulating soil loss rate in Ekbatan Dam watershed using experimental and statistical approaches. Int J Sediment Res 34: 226–239. https://doi.org/10.1016/j.ijsrc.2018.10.013 doi: 10.1016/j.ijsrc.2018.10.013
|
| [65] |
Amanollahi J, Ausati Sh (2020b). Validation of linear, nonlinear, and hybrid models for predicting particulate matter concentration in Tehran. Iran. Theor Appl Climatol 140: 709–717. https://doi.org/10.1007/s00704-020-03115-5 doi: 10.1007/s00704-020-03115-5
|
| [66] | Jorde K, Schneider M, Peter A, et al. (2001) Fuzzy based Models for the Evaluation of Fish Habitat Quality and Instream Flow Assessment. Proceedings of the 2001 International Symposium on Environmental Hydraulics Copyright © 2001, ISEH. |
| [67] |
Fukuda Sh, Hiramatsu K, Mori M (2006) Fuzzy neural network model for habitat prediction and HEP for habitat quality estimation focusing on 61 Japanese medaka (Oryzias latipes) in agricultural canals. Paddy Water Environ 4: 119–124. https://doi.org/10.1007/s10333-006-0039-5 doi: 10.1007/s10333-006-0039-5
|
| [68] |
Marsili-Libelli S, Giusti E, Nocita A (2012) A new instream flow assessment method based on fuzzy habitat suitability and large scale river modelling. Environ Modell Softw 41: 27–38. https://doi.org/10.1016/j.envsoft.2012.10.005 doi: 10.1016/j.envsoft.2012.10.005
|
| [69] |
Junga SH, Choib S-Uk (2015) Prediction of composite suitability index for physical habitatsimulations using the ANFIS method. Appl Soft Comput 34: 502–512. https://doi.org/10.1016/j.asoc.2015.05.028 doi: 10.1016/j.asoc.2015.05.028
|
| [70] |
Im D, Choi S-U, Choi B (2017) Physical habitat simulation for a fish community using the ANFIS method. Ecol Inform 43: 73–83. https://doi.org/10.1016/j.ecoinf.2017.09.001 doi: 10.1016/j.ecoinf.2017.09.001
|
| [71] | Elkiran G, Nourani V, Abba1 SI, et al. (2018) Artificial intelligence-based approaches for multi-station modelling of dissolve oxygen in river. Global J Environ Sci Manage 4: 439–450. |
Figures(8) / Tables(2)

Seiran Haghgoo, Jamil Amanollahi, Barzan Bahrami Kamangar, Shahryar Sorooshian. Decision models enhancing environmental flow sustainability: A strategic approach to water resource management[J]. AIMS Environmental Science, 2024, 11(6): 900-917. doi: 10.3934/environsci.2024045







 DownLoad:
DownLoad: