Citation: Angel Valverde, María González-Tirante, Marisol Medina-Sierra, Raúl Rivas, Ignacio Santa-Regina, José M. Igual. Culturable bacterial diversity from the chestnut (Castanea sativa Mill.) phyllosphere and antagonism against the fungi causing the chestnut blight and ink diseases[J]. AIMS Microbiology, 2017, 3(2): 293-314. doi: 10.3934/microbiol.2017.2.293

| [1] | Conedera M, Krebs P, Tinner W, et al. (2004) The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg Hist Archaeobot 13: 161–179. |
| [2] | Fernández-López J, Alía R (2003) Technical guidelines for genetic conservation and use for chestnut (Castanea sativa), Rome, Italy: EUFORGEN. |
| [3] |
Braga N, Rodrigues F, Beatriz M, et al. (2015) Castanea sativa by-products: A review on added value and sustainable application. Nat Prod Res 29: 1–18. doi: 10.1080/14786419.2014.955488

|
| [4] | Bounousa G (2014) Perspectives and future of the chestnut industry in Europe and all over the world. Acta Hortic 1043: 19–22. |
| [5] |
Gomes-Laranjo J, Araujo-Alves J, Ferreira-Cardoso J, et al. (2004) Effect of chestnut ink disease on photosynthetic performance. J Phytopathol 152: 138–144. doi: 10.1111/j.1439-0434.2004.00814.x
|
| [6] | Montenegro D, Aguin O, Sainz MJ, et al. (2008) Diversity of vegetative compatibility types, distribution of mating types and occurrence of hypovirulence of Cryphonectria parasitica in chestnut stands in NW Spain. For Ecol Manage 256: 973–980. |
| [7] |
Vettraino AM, Morel O, Perlerou C, et al. (2005) Occurrence and distribution of Phytophthora species in European chestnut stands, and their association with ink disease and crown decline. Eur J Plant Pathol 111: 169–180. doi: 10.1007/s10658-004-1882-0
|
| [8] |
Choupina AB, Estevinho L, Martins IM (2014) Scientifically advanced solutions for chestnut ink disease. Appl Microbiol Biotechnol 98: 3905–3909. doi: 10.1007/s00253-014-5654-2
|
| [9] | Martins LM, Oliveira MT, Abreu CG (1999) Soils and climatic characteristic of chestnut stands that differ on the presence of the ink disease. Acta Hortic 494: 447–449. |
| [10] |
Waldboth M, Oberhuber W (2009) Synergistic effect of drought and chestnut blight (Cryphonectria parasitica) on growth decline of European chestnut (Castanea sativa). Forest Pathol 39: 43–55. doi: 10.1111/j.1439-0329.2008.00562.x
|
| [11] |
Desprez-Loustau ML, Robin C, Reynaud G, et al. (2007) Simulating the effects of a climate-change scenario on the geographical range and activity of forest-pathogenic fungi. Can J Plant Pathol 29: 101–120. doi: 10.1080/07060660709507447
|
| [12] |
Vacher C, Hampe A, Porté AJ, et al. (2016) The phyllosphere: Microbial jungle at the plant-climate interface. Ann Rev Ecol Evol Syst 47: 1–24. doi: 10.1146/annurev-ecolsys-121415-032238
|
| [13] |
Vorholt JA (2012) Microbial life in the phyllosphere. Nat Rev Microbiol 10: 828–840. doi: 10.1038/nrmicro2910
|
| [14] |
Meyer KM, Leveau JHJ (2012) Microbiology of the phyllosphere: A playground for testing ecological concepts. Oecologia 168: 621–629. doi: 10.1007/s00442-011-2138-2
|
| [15] |
Heaton JC, Jones K (2008) Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: a review. J Appl Microbiol 104: 613–626. doi: 10.1111/j.1365-2672.2007.03587.x
|
| [16] |
Lindow SE, Leveau JH (2002) Phyllosphere microbiology. Curr Opin Biotechnol 13: 238–243. doi: 10.1016/S0958-1669(02)00313-0
|
| [17] |
Rico L, Ogaya R, Terradas J, et al. (2014) Community structures of N2-fixing bacteria associated with the phyllosphere of a Holm oak forest and their response to drought. Plant Biol 16: 586–593. doi: 10.1111/plb.12082
|
| [18] |
Furnkranz M, Wanek W, Richter A, et al. (2008) Nitrogen fixation by phyllosphere bacteria associated with higher plants and their colonizing epiphytes of a tropical lowland rainforest of Costa Rica. ISME J 2: 561–570. doi: 10.1038/ismej.2008.14
|
| [19] |
Sandhu A, Halverson LJ, Beattie GA (2007) Bacterial degradation of airborne phenol in the phyllosphere. Environ Microbiol 9: 383–392. doi: 10.1111/j.1462-2920.2006.01149.x
|
| [20] | Manching HC, Balint-Kurti PJ, Stapleton AE (2014) Southern leaf blight disease severity is correlated with decreased maize leaf epiphytic bacterial species richness and the phyllosphere bacterial diversity decline is enhanced by nitrogen fertilization. Front Plant Sci 5: 403. |
| [21] |
Müller T, Behrendt U, Ruppel S, et al. (2016) Fluorescent pseudomonads in the phyllosphere of wheat: potential antagonists against fungal pPhytopathogens. Curr Microbiol 72: 383–389. doi: 10.1007/s00284-015-0966-8
|
| [22] |
Ritpitakphong U, Falquet L, Vimoltust A, et al. (2016) The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol 210: 1033–1043. doi: 10.1111/nph.13808
|
| [23] | Mazinani Z, Zamani M, Sardari S (2017) Isolation and identification of phyllospheric bacteria possessing antimicrobial activity from Astragalus obtusifolius, Prosopis juliflora, Xanthium strumarium and Hippocrepis unisiliqousa. Avicenna J Med Biotechnol 9: 31–37. |
| [24] |
Krechel A, Faupel A, Hallmann J, et al. (2002) Potato-associated bacteria and their antagonistic potential towards plant-pathogenic fungi and the plant-parasitic nematode Meloidogyne incognita (Kofoid & White) Chitwood. Can J Microbiol 48: 772–786. doi: 10.1139/w02-071
|
| [25] |
Fravel DR (2005) Commercialization and implementation of biocontrol. Annu Rev Phytopathol 43: 337–359. doi: 10.1146/annurev.phyto.43.032904.092924
|
| [26] |
Yadav RKP, Karamanoli K, Vokou D (2005) Bacterial colonization of the phyllosphere of Mediterranean perennial species as influenced by leaf structural and chemical features. Microb Ecol 50: 185–196. doi: 10.1007/s00248-004-0171-y
|
| [27] |
Peñuelas J, Rico L, Ogaya R, et al. (2012) Summer season and long-term drought increase the richness of bacteria and fungi in the foliar phyllosphere of Quercus ilex in a mixed Mediterranean forest. Plant Biol 14: 565–575. doi: 10.1111/j.1438-8677.2011.00532.x
|
| [28] | Valverde A, Igual JM, Regina IS, et al. (2005) Preliminary diversity studies of culturable phyllosphere bacteria on chestnut (Castanea sativa). Acta Hortic 693: 263–270. |
| [29] | Salazar-Iglesias S (2008) Estudio de los procesos ecológicos para el desarrollo sostenible del castaño (Castanea sativa Mill.) de la Sierra de Francia (PhD dissertation). Salamanca, Spain: University of Salamanca. |
| [30] |
Jacobs JL, Sundin GW (2001) Effect of solar UV-B radiation on a phyllosphere bacterial community. Appl Environ Microbiol 67: 5488–5496. doi: 10.1128/AEM.67.12.5488-5496.2001
|
| [31] |
Rivas R, Velazquez E, Valverde A, et al. (2001) A two primers random amplified polymorphic DNA procedure to obtain polymerase chain reaction fingerprints of bacterial species. Electrophoresis 22: 1086–1089. doi: 10.1002/1522-2683()22:6<1086::AID-ELPS1086>3.0.CO;2-6
|
| [32] |
Valverde A, Igual JM, Peix A, et al. (2006) Rhizobium lusitanum sp nov a bacterium that nodulates Phaseolus vulgaris. Int J Syst Evol Microbiol 56: 2631–2637. doi: 10.1099/ijs.0.64402-0
|
| [33] |
Altschul SF, Madden TL, Schäffer AA, et al. (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. doi: 10.1093/nar/25.17.3389
|
| [34] |
Pearson WR, Lipman DJ (1988) Improved tools for biological sequence comparison. Proc Natl Acad Sci USA 85: 2444–2448. doi: 10.1073/pnas.85.8.2444
|
| [35] | Yoon SH, Ha SM, Kwon S, et al. (2017) Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int J Syst Evol Microbiol (in press). |
| [36] |
Thompson JD, Gibson TJ, Plewniak F, et al. (1997) The CLUSTAL X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25: 4876–4882. doi: 10.1093/nar/25.24.4876
|
| [37] | Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406–425. |
| [38] |
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16: 111–120. doi: 10.1007/BF01731581
|
| [39] |
Tamura K, Dudley J, Nei M, et al. (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24: 1596–1599. doi: 10.1093/molbev/msm092
|
| [40] |
Ansari MA, Tirry L, Moens M (2005) Antagonism between entomopathogenic fungi and bacterial symbionts of entomopathogenic nematodes. Biocontrol 50: 465–475. doi: 10.1007/s10526-004-5524-4
|
| [41] | Bric JM, Bostock RM, Silverstone SE (1991) Rapid in situ assay for indoleacetic-acid production by bacteria immobilized on a nitrocellulose membrane. Appl Environ Microbiol 57: 535–538. |
| [42] | Chao A, Shen TJ (2003) Program SPADE (Species Prediction and Diversity Estimation), Program and User's Guide. Available from: http://chao.stat.nthu.edu.tw/word press/software_download/. |
| [43] |
Colwell RK, Elsensohn JE (2014) EstimateS turns 20: Statistical estimation of species richness and shared species from samples, with non-parametric extrapolation. Ecography 37: 609–613. doi: 10.1111/ecog.00814
|
| [44] | Achtman M, Wagner M (2008) Microbial diversity and the genetic nature of microbial species. Nat Rev Microbiol 6: 431–440. |
| [45] | Stackebrandt E, Frederiksen W, Garrity GM, et al. (2002) Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int J Syst Evol Microbiol 52: 1043–1047. |
| [46] |
Rivas R, Velázquez E, Palomo JL, et al. (2002) Rapid identification of Clavibacter michiganensis subspecies sepedonicus using two primers random amplified polymorphic DNA (TP-RAPD) fingerprints. Eur J Plant Pathol 108: 179–184. doi: 10.1023/A:1015044911913
|
| [47] | Valverde A, Peix A, Rivas R, et al. (2008) Paenibacillus castaneae sp nov., isolated from the phyllosphere of Castanea sativa Miller. Int J Syst Evol Microbiol 58: 2560–2564. |
| [48] |
Velázquez E, Rojas M, Lorite MJ, et al. (2008) Genetic diversity of endophytic bacteria which could be find in the apoplastic sap of the medullary parenchym of the stem of healthy sugarcane plants. J Basic Microbiol 48: 118–124. doi: 10.1002/jobm.200700161
|
| [49] |
Huey B, Hall J (1989) Hypervariable DNA fingerprinting in Escherichia coli: minisatellite probe from bacteriophage M13. J Bacteriol 171: 2528–2532. doi: 10.1128/jb.171.5.2528-2532.1989
|
| [50] |
Velázquez E, Valverde A, Rivas R, et al. (2010) Strains nodulating Lupinus albus on different continents belong to several new chromosomal and symbiotic lineages within Bradyrhizobium. Antonie Van Leeuwenhoek 97: 363–376. doi: 10.1007/s10482-010-9415-7
|
| [51] |
Laforest-Lapointe I, Messier C, Kembel SW (2016) Tree phyllosphere bacterial communities: Exploring the magnitude of intra- and inter-individual variation among host species. PeerJ 4: e2367. doi: 10.7717/peerj.2367
|
| [52] |
Becker R, Behrendt U, Hommel B, et al. (2008) Effects of transgenic fructan-producing potatoes on the community structure of rhizosphere and phyllosphere bacteria. FEMS Microbiol Ecol 66: 411–425. doi: 10.1111/j.1574-6941.2008.00562.x
|
| [53] |
Redford AJ, Bowers RM, Knight R, et al. (2010) The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ Microbiol 12: 2885–2893. doi: 10.1111/j.1462-2920.2010.02258.x
|
| [54] | Kumar J, Babele PK, Singh D, et al. (2016) UV-B radiation stress causes alterations in whole cell protein profile and expression of certain genes in the rice phyllospheric bacterium Enterobacter cloacae. Front Microbiol 7: 1440. |
| [55] |
Kadivar H, Stapleton AE (2003) Ultraviolet radiation alters maize phyllosphere bacterial diversity. Microb Ecol 45: 353–361. doi: 10.1007/s00248-002-1065-5
|
| [56] |
De la Vega UP, Rettberg P, Douki T, et al. (2005) Sensitivity to polychromatic UV-radiation of strains of Deinococcus radiodurans differing in their DNA repair capacity. Int J Radiat Biol 81: 601–611. doi: 10.1080/09553000500309374
|
| [57] |
Jacobs JL, Carroll TL, Sundin GW (2005) The role of pigmentation, ultraviolet radiation tolerance, and leaf colonization strategies in the epiphytic survival of phyllosphere bacteria. Microb Ecol 49: 104–113. doi: 10.1007/s00248-003-1061-4
|
| [58] |
Cazorla FM, Codina JC, Abad C, et al. (2008) 62-kb plasmids harboring rulAB homologues confer UV-tolerance and epiphytic fitness to Pseudomonas syringae pv. syringae mango isolates. Microb Ecol 56: 283–291. doi: 10.1007/s00248-007-9346-7
|
| [59] |
Martirosyan V, Unc A, Miller G, et al. (2016) Desert Perennial Shrubs Shape the Microbial-Community Miscellany in Laimosphere and Phyllosphere Space. Microb Ecol 72: 659–668. doi: 10.1007/s00248-016-0822-9
|
| [60] |
Ueda Y, Frindte K, Knief C, et al. (2016) Effects of elevated tropospheric ozone concentration on the bacterial community in the phyllosphere and rhizoplane of rice. PLoS One 11: e0163178. doi: 10.1371/journal.pone.0163178
|
| [61] |
Ren G, Zhu C, Alam MS, et al. (2015) Response of soil, leaf endosphere and phyllosphere bacterial communities to elevated CO2 and soil temperature in a rice paddy. Plant Soil 392: 27–44. doi: 10.1007/s11104-015-2503-8
|
| [62] | Fonseca-García C, Coleman-Derr D, Garrido E, et al. (2016) The cacti microbiome: interplay between habitat-filtering and host-specificity. Front Microbiol 7: 150. |
| [63] |
Kembel SW, O'Connor TK, Arnold HK, et al. (2014) Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc Natl Acad Sci USA 111: 13715–13720. doi: 10.1073/pnas.1216057111
|
| [64] |
Dees MW, Lysøe E, Nordskog B, et al. (2015) Bacterial communities associated with surfaces of leafy greens: Shift in composition and decrease in richness over time. Appl Environ Microbiol 81: 1530–1539. doi: 10.1128/AEM.03470-14
|
| [65] |
Müler T, Müller M, Behrendt U, et al. (2003) Diversity of culturable phyllosphere bacteria on beech and oak: the effects of lepidopterous larvae. Microbiol Res 158: 291–297. doi: 10.1078/0944-5013-00207
|
| [66] |
Lambais MR, Barrera SE, Santos EC, et al. (2017) Phyllosphere metaproteomes of trees from the Brazilian Atlantic forest show high levels of functional redundancy. Microb Ecol 73: 123–134. doi: 10.1007/s00248-016-0878-6
|
| [67] | Behrendt U, Ulrich A, Schumann P (2003) Fluorescent pseudomonads associated with the phyllosphere of grasses; Pseudomonas trivialis sp nov., Pseudomonas poae sp nov and Pseudomonas congelans sp nov. Int J Syst Evol Microbiol 53: 1461–1469. |
| [68] | Behrendt U, Ulrich A, Schumann P, et al. (2002) Diversity of grass-associated Microbacteriaceae isolated from the phyllosphere and litter layer after mulching the sward; polyphasic characterization of Subtercola pratensis sp nov., Curtobacterium herbarum sp nov and Plantibacter flavus gen. nov., sp nov. Int J Syst Evol Microbiol 52: 1441–1454. |
| [69] |
Lambais MR, Crowley DE, Cury JC, et al. (2006) Bacterial diversity in tree canopies of the Atlantic forest. Science 312: 1917. doi: 10.1126/science.1124696
|
| [70] |
Rivas R, Garcia-Fraile P, Mateos PF, et al. (2007) Characterization of xylanolytic bacteria present in the bract phyllosphere of the date palm Phoenix dactylifera. Lett Appl Microbiol 44: 181–187. doi: 10.1111/j.1472-765X.2006.02050.x
|
| [71] |
Yang CH, Crowley DE, Borneman J, et al. (2001) Microbial phyllosphere populations are more complex than previously realized. Proc Natl Acad Sci USA 98: 3889–3894. doi: 10.1073/pnas.051633898
|
| [72] |
Lindow SE, Brandl MT (2003) Microbiology of the phyllosphere. Appl Environ Microbiol 69: 1875–1883. doi: 10.1128/AEM.69.4.1875-1883.2003
|
| [73] |
Jackson EF, Echlin HL, Jackson CR (2006) Changes in the phyllosphere community of the resurrection fern, Polypodium polypodioides, associated with rainfall and wetting. FEMS Microbiol Ecol 58: 236–246. doi: 10.1111/j.1574-6941.2006.00152.x
|
| [74] | Brady CL, Cleenwerck I, Denman S, et al. (2012) Proposal to reclassify Brenneria quercina (Hildebrand and Schroth 1967) Hauben et al. 1999 into a new genus, Lonsdalea gen. nov., as Lonsdalea quercina comb. nov., descriptions of Lonsdalea quercina subsp. quercina comb. nov., Lonsdalea quercina subsp. iberica subsp. nov. and Lonsdalea quercina subsp. britannica subsp. nov. Int J Syst Evol Microbiol 62: 1592–1602. |
| [75] |
Enya J, Shinohara H, Yoshida S, et al. (2007) Culturable leaf-associated bacteria on tomato plants and their potential as biological control agents. Microb Ecol 53: 524–536. doi: 10.1007/s00248-006-9085-1
|
| [76] |
Liu CH, Chen X, Liu TT, et al. (2007) Study of the antifungal activity of Acinetobacter baumannii LCH001 in vitro and identification of its antifungal components. Appl Microbiol Biotechnol 76: 459–466. doi: 10.1007/s00253-007-1010-0
|
| [77] |
Liu X, Jia J, Atkinson S, et al. (2010) Biocontrol potential of an endophytic Serratia sp G3 and its mode of action. World J Microbiol Biotechnol 26: 1465–1471. doi: 10.1007/s11274-010-0321-y
|
| [78] | Bedini S, Bagnoli G, Sbrana C, et al. (1999) Pseudomonads isolated from within fruit bodies of Tuber borchii are capable of producing biological control or phytostimulatory compounds in pure culture. Symbiosis 26: 223–236. |
| [79] |
Wilhelm E, Arthofer W, Schafleitner R, et al. (1998) Bacillus subtilis an endophyte of chestnut (Castanea sativa) as antagonist against chestnut blight (Cryphonectria parasitica). Plant Cell Tissue Organ Cult 52: 105–108. doi: 10.1023/A:1005917906769
|
| [80] | Aravind R, Kumar A, Eapen SJ, et al. (2009) Endophytic bacterial flora in root and stem tissues of black pepper (Piper nigrum L.) genotype: isolation, identification and evaluation against Phytophthora capsici. Lett Appl Microbiol 48: 58–64. |
| [81] |
Cirvilleri G, Spina S, Iacona C, et al. (2008) Study of rhizosphere and phyllosphere bacterial community and resistance to bacterial canker in genetically engineered phytochrome A cherry plants. J Plant Physiol 165: 1107–1119. doi: 10.1016/j.jplph.2008.01.009
|
| [82] |
Sharma A, Wray V, Johri BN (2007) Rhizosphere Pseudomonas sp strains reduce occurrence of pre- and post-emergence damping-off in chile and tomato in Central Himalayan region. Arch Microbiol 187: 321–335. doi: 10.1007/s00203-006-0197-2
|
| [83] |
Timmusk S, van West P, Gow NAR, et al. (2009) Paenibacillus polymyxa antagonizes oomycete plant pathogens Phytophthora palmivora and Pythium aphanidermatum. J Appl Microbiol 106: 1473–1481. doi: 10.1111/j.1365-2672.2009.04123.x
|
| [84] | Verma VC, Gond SK, Kumar A, et al. (2009) Endophytic Actinomycetes from Azadirachta indica A. Juss.: isolation, diversity, and anti-microbial activity. Microb Ecol 57: 749–756. |
| [85] |
Weinert N, Meincke R, Gottwald C, et al. (2010) Bacterial diversity on the surface of potato tubers in soil and the influence of the plant genotype. FEMS Microbiol Ecol 74: 114–123. doi: 10.1111/j.1574-6941.2010.00936.x
|
| [86] |
Biondi N, Piccardi R, Margheri MC, et al. (2004) Evaluation of Nostoc strain ATCC 53789 as a potential source of natural pesticides. Appl Environ Microbiol 70: 3313–3320. doi: 10.1128/AEM.70.6.3313-3320.2004
|
| [87] |
El-Tarabily KA, Sivasithamparam K (2006) Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biol Biochem 38: 1505–1520. doi: 10.1016/j.soilbio.2005.12.017
|
| [88] |
Kavroulakis N, Ntougias S, Besi MI, et al. (2010) Antagonistic bacteria of composted agro-industrial residues exhibit antibiosis against soil-borne fungal plant pathogens and protection of tomato plants from Fusarium oxysporum f.sp radicis-lycopersici. Plant Soil 333: 233–247. doi: 10.1007/s11104-010-0338-x
|
| [89] |
Glandorf DCM, Verheggen P, Jansen T, et al. (2001) Effect of genetically modified Pseudomonas putida WCS358r on the fungal rhizosphere microflora of field-grown wheat. Appl Environ Microbiol 67: 3371–3378. doi: 10.1128/AEM.67.8.3371-3378.2001
|
| [90] |
Khan MR, Fischer S, Egan D, et al. (2006) Biological control of fusarium seedling blight disease of wheat and barley. Phytopathology 96: 386–394. doi: 10.1094/PHYTO-96-0386
|
| [91] |
Lee SW, Cooksey DA (2000) Genes expressed in Pseudomonas putida during colonization of a plant-pathogenic fungus. Appl Environ Microbiol 66: 2764–2772. doi: 10.1128/AEM.66.7.2764-2772.2000
|
| [92] |
Brandl MT, Quinones B, Lindow SE (2001) Heterogeneous transcription of an indoleacetic acid biosynthetic gene in Erwinia herbicola on plant surfaces. Proc Natl Acad Sci USA 98: 3454–3459. doi: 10.1073/pnas.061014498
|
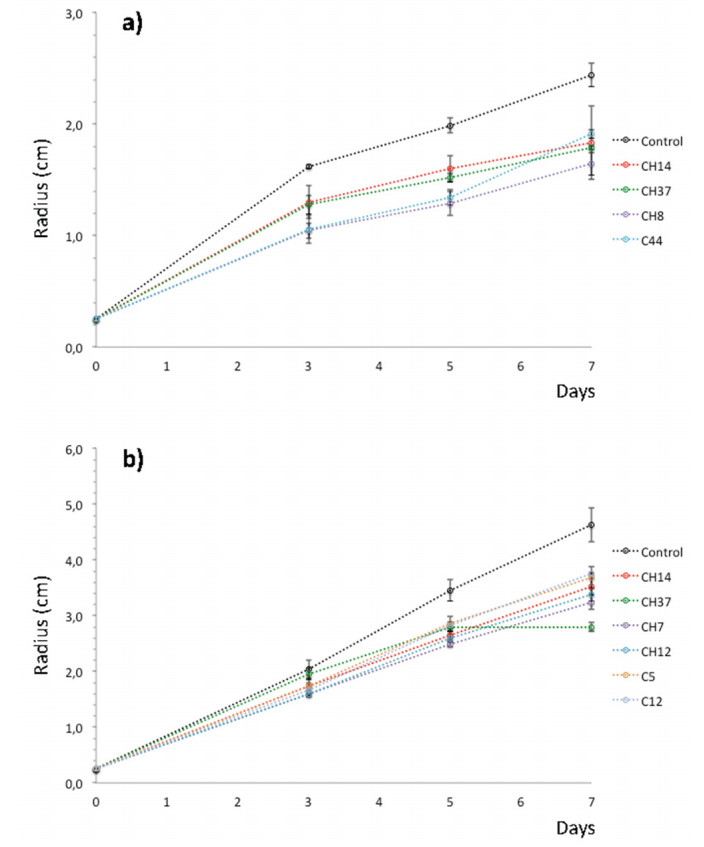
Figures(5) / Tables(3)

Angel Valverde, María González-Tirante, Marisol Medina-Sierra, Raúl Rivas, Ignacio Santa-Regina, José M. Igual. Culturable bacterial diversity from the chestnut (Castanea sativa Mill.) phyllosphere and antagonism against the fungi causing the chestnut blight and ink diseases[J]. AIMS Microbiology, 2017, 3(2): 293-314. doi: 10.3934/microbiol.2017.2.293







 DownLoad:
DownLoad: