Citation: Camilo Muñoz, Fernando G. Fermoso, Mariella Rivas, Juan M. Gonzalez. Hydrolytic enzyme activity enhanced by Barium supplementation[J]. AIMS Microbiology, 2016, 2(4): 402-411. doi: 10.3934/microbiol.2016.4.402

| [1] | Sexton S, Zilberman D, Rajagopal D, et al. (2009) The role of biotechnology in a sustainable biofuel future. AgBioForum 12: 130–140. |
| [2] | Bhalla A, Bansal N, Kumar S, et al. (2013) Improved lignocellulose conversion to biofuels with thermophilic bacteria and thermostable enzymes. Bioresour Technol 128: 751–759. |
| [3] | Horn SJ, Vaaje-Kolstad G, Westereng B, et al. (2012) Novel enzymes for the degradation of cellulose. Biotechnol Biofuels 5: 89–93. |
| [4] | Shubert R, Schellnhuber HJ, Buchmann N, et al. (2010) Future bioenergy and sustainable land use. German Advisory Council on Global Change. London: Earthscan. |
| [5] | Kein-Marcuschamer D, Oleskowicz-Popiel P, Simmons PA, et al. (2012) The challenge of enzyme cost in the production of lignocellulosic biofuels. Biotechnol Bioeng 109: 1083–1087. |
| [6] | Jeffries TW (1990) Biodegradation of lignin-carbohydrate complexes. Biodegradation 1: 163–176. |
| [7] | Fulekar MH (2010) Environmental Biotechnology, Boca raton, Florida: CRC Press, 308. |
| [8] | Li Q, Du W, Liu D (2008) Perspectives of microbial oils for biodiesel production. Appl Microbiol Biotechnol 80: 749–756. |
| [9] | Zandvoort MH, van Hullebush ED, Fermoso FG, et al. (2006) Trace metals in anaerobic granular sludge reactors: bioavailability and dosing strategies. Eng Life Sci 6: 293–301. |
| [10] | Portillo MC, Reina M, Serrano L, et al. (2008) Role of specific microbial communities in the bioavailability of iron in Doñana National Park. Environ Geochem Health 30: 165–170. |
| [11] |
Rho M, Tang H, Ye Y (2010) FragGeneScan: predicting genes in short and error-prone reads. Nucl Acids Res 38: e191. doi: 10.1093/nar/gkq747

|
| [12] | Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl Acids Res 22: 4673–4680. |
| [13] | Zhu Y, Liu G, Li H, et al. (2012) Cloning and characterization of a thermostable carboxylesterase from inshore hot spring thermophile Geobacillussp. ZH1. Acta Oceanol Sin 31: 117–126. |
| [14] | Asoodeh A, Ghanbari T (2013) Characterization of an extracellular thermophilic alkaline esterase produced by Bacillus subtilis DR8806. J Mol Catal B 85–86: 49–55. |
| [15] | Brault G, Shareck F, Hurtubise Y, et al. (2012) Isolation and characterization of EstC, a new cold-active esterase from Streptomyces coelicolor A3(2). PloS One 7(3): e32041. |
| [16] | Mohamed YM, Ghazy MA, Sayed A, et al. (2013) Isolation and characterization of a heavy metal-resistant, thermophilic esterase from a Red Sea Brine Pool. Scientific Rep 3: 3358–3358. |
| [17] | Andrade JP, da Rocha Bispo AS, Marbach PAS, et al. (2011) Production and partial characterization of cellulases from Trichoderma sp. IS-05 isolated from sandy coastal plains of Northeast Brazil. Enz Res 167248: 1–7. |
| [18] | Ling L, Kan X, Hao Y, et al. (2012) Characterization of extracellular cellulose-degrading enzymes from Bacillus thuringiensis strains. Electronic J Biotechnol 15: 717–3458. |
| [19] | Buchholz K, Kasche V, Bornscheuer UT (2012) Biocatalysts and enzyme technology, 2 Eds., Hoboken, New Jersey: Wiley-Blackwell. |
| [20] | Madigan MT, Martinko JM, Parker J (2003) Brock Biology of microorganisms. New Jersey: Prentice-Hall Inc. |
| [21] | De Azeredo LAI, Freire DMG, Soares RMA, et al. (2004) Production and partial characterization of thermophilic proteases from Streptomyces sp. Isolated from Brazilian Cerrado soil. Enz Microb Technol 34: 354–358. |
| [22] | Jayakumar R, Jayashree S, Annapurna B, et al. (2012) Characterization of thermophilic serine alkaline protease from an alkaliphilic strain Bacillus pumilus MCAS8 and its applications. Appl Biochem Biotechnol 168: 1849–1866. |
| [23] | Deb P, Talukdar SA, Mohsina K, et al. (2013) Production and partial characterization of extracellular amylase enzyme from Bacillus amyloliquefaciens P-001. SpringerPlus 2: 1–12. |
| [24] | Kim DW, Jang YH, Kim CS, et al. (2011) Effect of metal ions on the degradation and adsorption of two cellobiohydrolases on microcrystalline cellulose. Bull Korean Chem Soc 22: 716–720. |
| [25] | Shanker AK (2008) Mode of action and toxicity of trace metals. In: Prasad MNV, editor, Trace elements as contaminants and nutrients: consequences in ecosystems and human health. Hoboken, New Jersey: John Wiley & Sons Inc. |
| [26] | Kresse R, Baudis U, Jäger P, et al. (2007) Barium and barium compounds. Ullmann’s Encyclopedia of Industrial Chemistry. Boca Raton, Florida: CRC Press. |
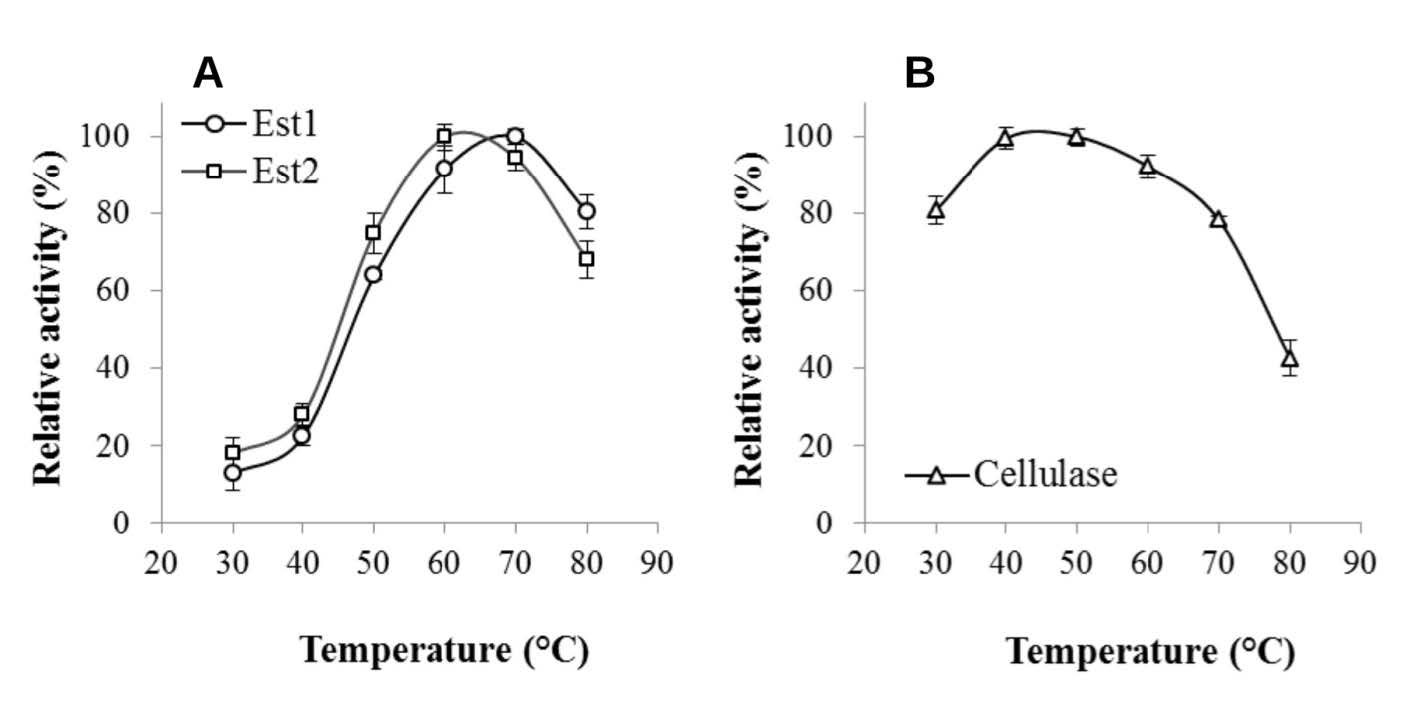
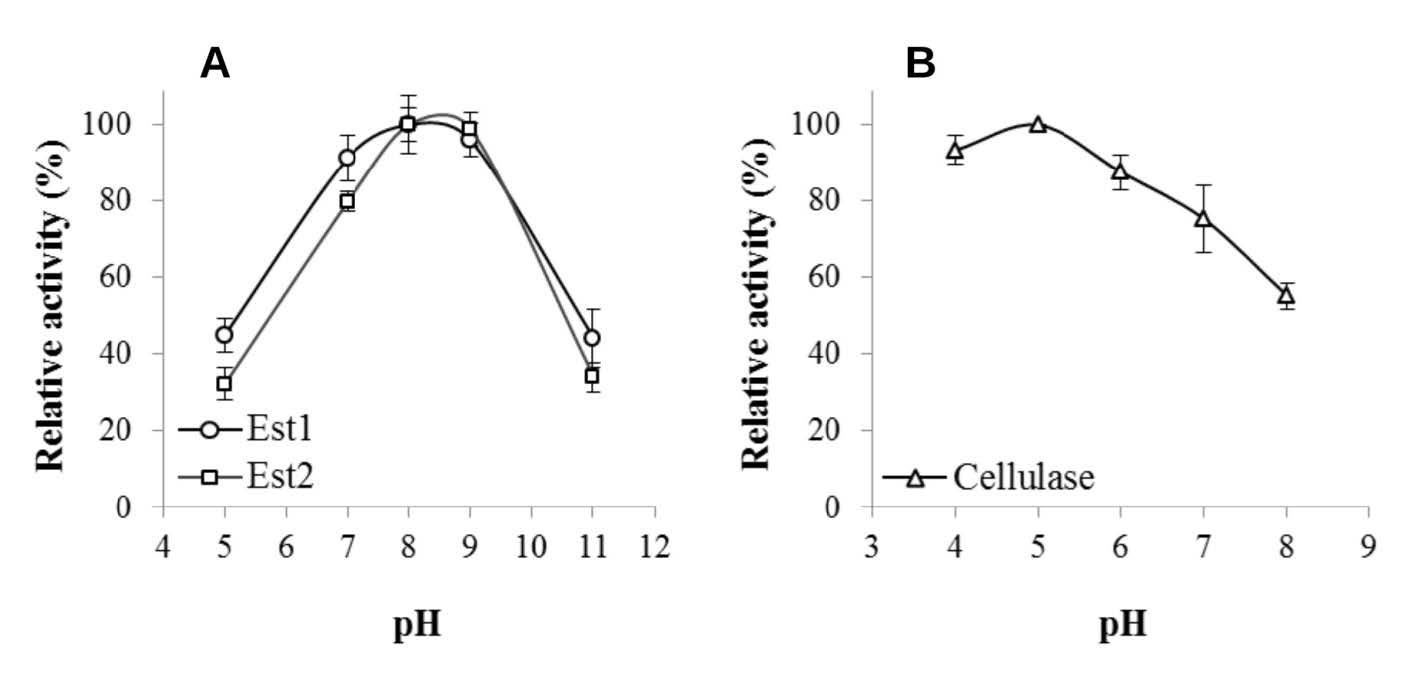
Figures(2) / Tables(1)

Camilo Muñoz, Fernando G. Fermoso, Mariella Rivas, Juan M. Gonzalez. Hydrolytic enzyme activity enhanced by Barium supplementation[J]. AIMS Microbiology, 2016, 2(4): 402-411. doi: 10.3934/microbiol.2016.4.402







 DownLoad:
DownLoad: