Citation: K. Wayne Forsythe, Chris H. Marvin, Danielle E. Mitchell, Joseph M. Aversa, Stephen J. Swales, Debbie A. Burniston, James P. Watt, Daniel J. Jakubek, Meghan H. McHenry, Richard R. Shaker. Utilization of bathymetry data to examine lead sediment contamination distributions in Lake Ontario[J]. AIMS Environmental Science, 2016, 3(3): 347-361. doi: 10.3934/environsci.2016.3.347

| [1] | U.S. Environmental Protection Agency and Government of Canada (USEPAGC), (1995) The Great Lakes—An Environmental Atlas and Resource Book (3rd Edition), Chicago: Great Lakes National Program Office. |
| [2] | Great Lakes Information Network (GLIN), 2004. Great Lakes Facts and Figures. Great Lakes Information Network. Available from: http://www.great-lakes.net/lakes/ref/lakefact.html |
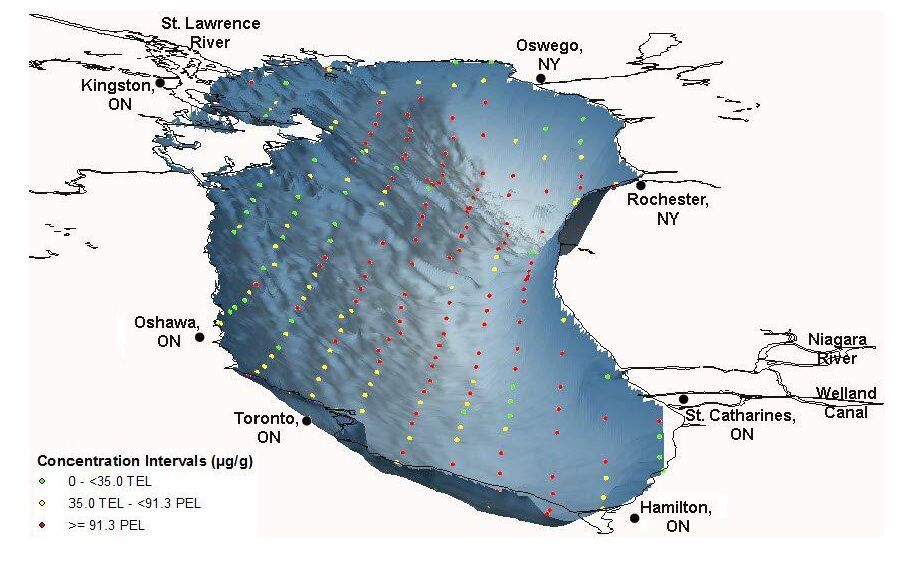
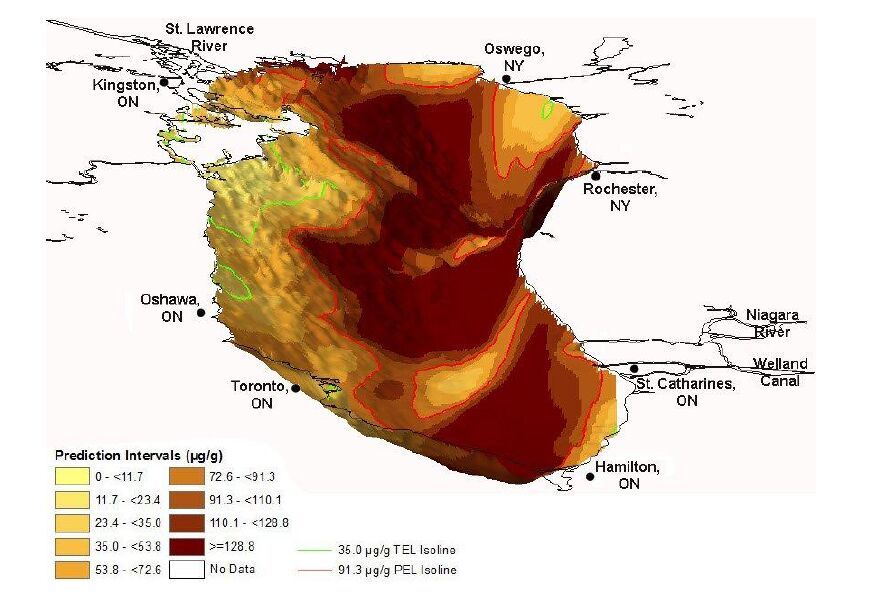
| [3] | Crane J, MacDonald DD (2003) Applications of Numerical Sediment Quality Targets for Assessing Sediment Quality Conditions in a US Great Lakes Area of Concern. Environ Manage 32: 128-140. |
| [4] | Canadian Council of Ministers of the Environment (CCME), 1999. Canadian Environmental Quality Guidelines. Winnipeg, MB: Canadian Council of Ministers of the Environment. |
| [5] | Forsythe KW, Marvin CH, Valancius CJ, et al. (2016) Using Geovisualization to Assess Lead Sediment Contamination in Lake St. Clair. The Canadian Geographer/Le Géographe canadien 60: 149-158. |
| [6] | Newell RG, Rogers K, The U.S. Experience with the phasedown of lead in gasoline, 2003. Available from: http://web.mit.edu/ckolstad/www/Newell.pdf |
| [7] | Currie E (1994) Contaminated Sediment Removal Program Environment Canada—Great Lakes Cleanup Fund. |
| [8] | Forsythe KW, Watt JP (2006) Using Geovisualization to Assess Sediment Contamination in the “Sixth” Great Lake. In: Strobl J, Blaschke T, Griesebner G (Eds), Proceedings of the 18th Symposium for Applied Geographic Information Processing, AGIT XVIII, Salzburg, Austria, July 5-7, 2006, Herbert Wichmann Verlag, Hüthig GmbH & Co. KG: Heidelberg, Germany, 161-170. |
| [9] | Forsythe KW, Marvin CH, Valancius CJ, et al. (2016) Geovisualization of Mercury Contamination in Lake St. Clair Sediments. J Mar Sci Eng 4: 19. |
| [10] | Forsythe KW, Dennis M, Marvin CH (2004) Comparison of Mercury and Lead Sediment Concentrations in Lake Ontario (1968–1998) and Lake Erie (1971-1997/98) using a GIS-based Kriging Approach. Water Qual Res J Can 39: 190-206. |
| [11] | Jakubek DJ, Forsythe KW (2004) A GIS-based kriging approach for assessing Lake Ontario sediment contamination. Great Lakes Geographer 11: 1-14. |
| [12] | Forsythe KW, Marvin CH (2005) Analyzing the Spatial Distribution of Sediment Contamination in the Lower Great Lakes. Water Qual Res J Can 40: 389-401. |
| [13] | Gewurtz SB, Helm PA, Waltho J, et al. (2007) Spatial distributions and temporal trends in sediment contamination in Lake St. Clair. J Great Lakes Res 33: 668-685. |
| [14] | Gewurtz SB, Shen L, Helm PA, et al. (2008) Spatial distribution of legacy contaminants in sediments of Lakes Huron and Superior. J Great Lakes Res 34: 153-168. |
| [15] | Forsythe KW, Marvin CH (2009) Assessing Historical versus Contemporary Mercury and Lead Contamination in Lake Huron Sediments. Aquat Ecosyst Health 12: 101-109. |
| [16] |
Forsythe KW, Paudel K, Marvin CH (2010) Geospatial analysis of zinc contamination in Lake Ontario sediments. J Environ Inform 16: 1-10. doi: 10.3808/jei.201000172

|
| [17] | Gawedzki A, Forsythe KW (2012) Assessing anthracene and arsenic contamination within Buffalo River sediments. Int J Ecol 2012: Article ID 496740. |
| [18] | Forsythe KW, Irvine KN, Atkinson DM, et al. (2015) Assessing lead contamination in Buffalo River sediments. J Environ Inform 26: 106-111. |
| [19] | Johnston K, Ver Hoef J, Krivoruchko K, et al. (2001) Using ArcGIS geostatistical analyst. Redlands, CA: Environmental Systems Research Institute. |
| [20] | Isaaks EH, Srivastava MR (1989) An Introduction to Applied Geostatistics. New York, NY: Oxford University Press. |
| [21] | National Oceanographic and Atmospheric Administration (NOAA), Great Lakes bathymetry. 2014. Available from: http://www.ngdc.noaa.gov/mgg/greatlakes/greatlakes.html. |
| [22] | Great Lakes Information Network (GLIN), 2004. Lake Ontario Facts and Figures. Great Lakes Information Network. Available from: http://www.great-lakes.net/lakes/ref/ontfact.html |
| [23] |
Marvin CH, Charlton MN, Reiner EJ, et al. (2002) Surficial sediment contamination in Lakes Erie and Ontario: A comparative analysis. J Great Lakes Res 28: 437-450. doi: 10.1016/S0380-1330(02)70596-0
|
| [24] | Marvin CH, Painter S, Rossmann R (2004) Spatial and temporal patterns in mercury contamination in sediments of the Laurentian Great Lakes. Environ Res 95: 351-362. |
| [25] |
Marvin CH, Painter S, Charlton MN, et al. (2004) Trends in spatial and temporal levels of persistent organic pollutants in Lake Erie sediments. Chemosphere 54: 33-40. doi: 10.1016/S0045-6535(03)00660-X
|
| [26] |
Dunn RJK, Zigic S, Burling M, et al. (2015) Hydrodynamic and Sediment Modelling within a Macro Tidal Estuary: Port Curtis Estuary, Australia. J Mar Sci Eng 3: 720-744. doi: 10.3390/jmse3030720
|
| [27] |
Hessl A, Miller J, Kernan J, et al. (2007) Mapping Paleo-Fire Boundaries from Binary Point Data: comparing Interpolation Methods. The Professional Geographer 59: 87-104. doi: 10.1111/j.1467-9272.2007.00593.x
|
| [28] |
Niu Y, Jiao W, Yu H, et al. (2015) Spatial Evaluation of Heavy Metals Concentrations in the Surface Sediment of Taihu Lake. Int J Environ Res Public Health 12: 15028-15039. doi: 10.3390/ijerph121214966
|
| [29] |
Ouyang Y, Higman J, Campbell D, et al. (2003) Three-Dimensional Kriging Analysis of Sediment Mercury Distribution: A Case Study. J Am Water Resour Assoc 39: 689-702. doi: 10.1111/j.1752-1688.2003.tb03685.x
|
| [30] |
Marvin CH, Charlton MN, Stern GA, et al. (2003) Spatial and temporal trends in sediment contamination in Lake Ontario. J Great Lakes Res 29: 317-331. doi: 10.1016/S0380-1330(03)70437-7
|
| [31] |
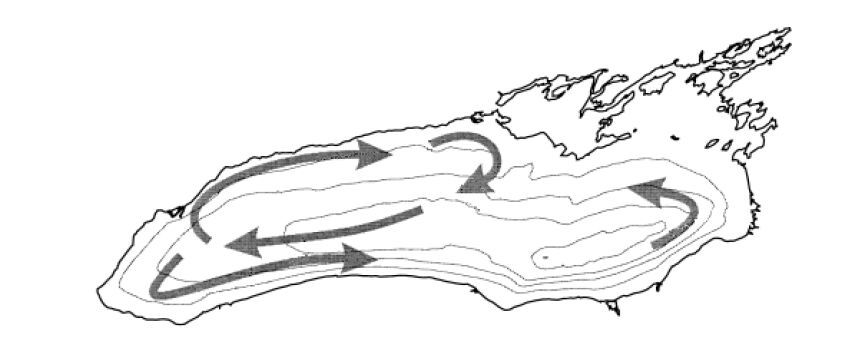
Beletsky D, Saylor JH, Schwab DJ (1999). Mean circulation in the Great Lakes. J Great Lakes Res 25: 78-93. doi: 10.1016/S0380-1330(99)70718-5
|
| [32] | Environment Canada, Environment Canada’s gasoline regulations, 2009. Available from: http://www.ec.gc.ca/lcpe-cepa/default.asp?lang¼En&n¼54FE5535-1&wsdoc¼8E3C2E9B-38A8-461A-8EC3-C3AA3B1FD585. |
| [33] |
Marvin CH, Charlton MN, Stern GA, et al. (2003) Spatial and temporal trends in sediment contamination in Lake Ontario. J Great Lakes Res 29: 317-331. doi: 10.1016/S0380-1330(03)70437-7
|
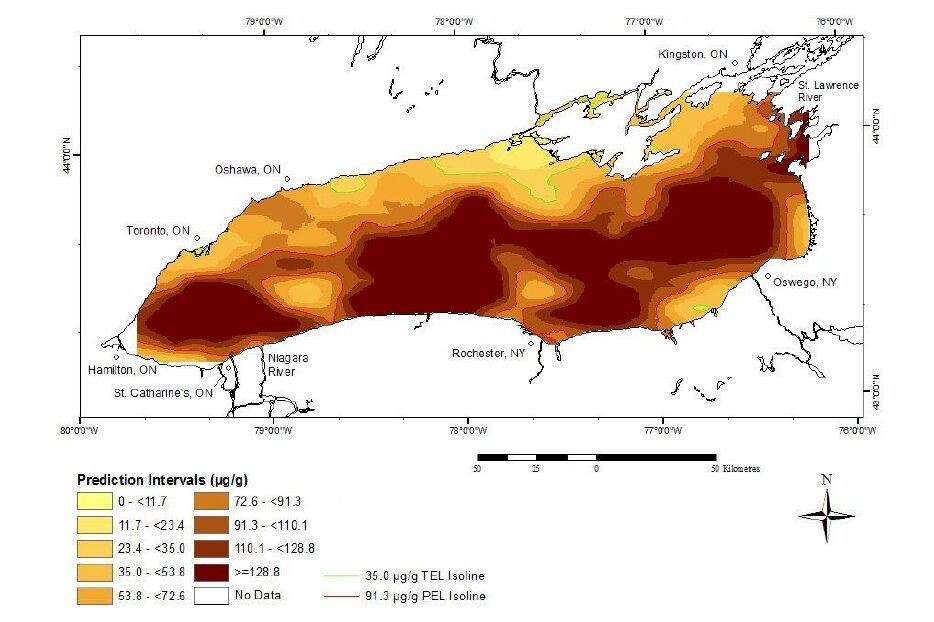
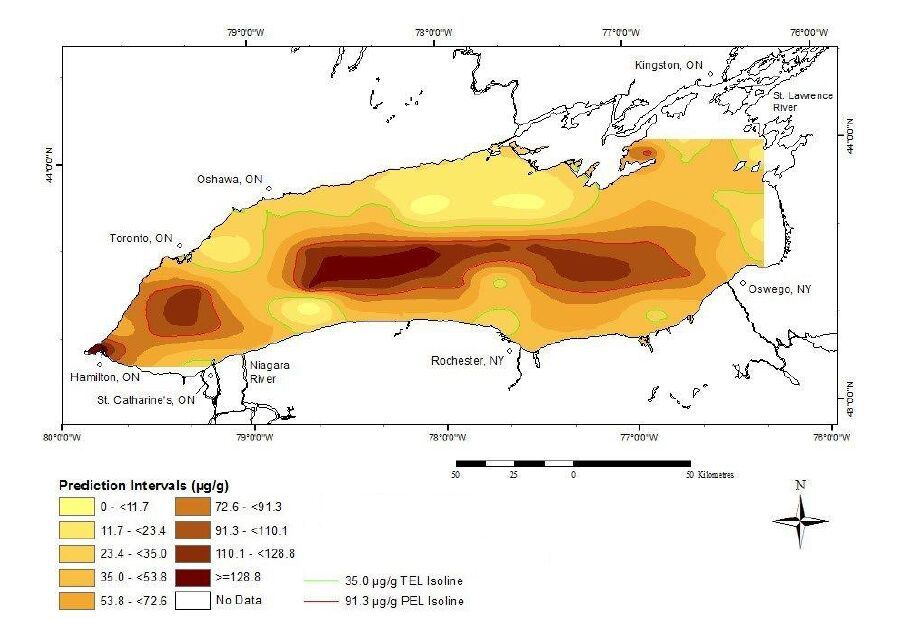
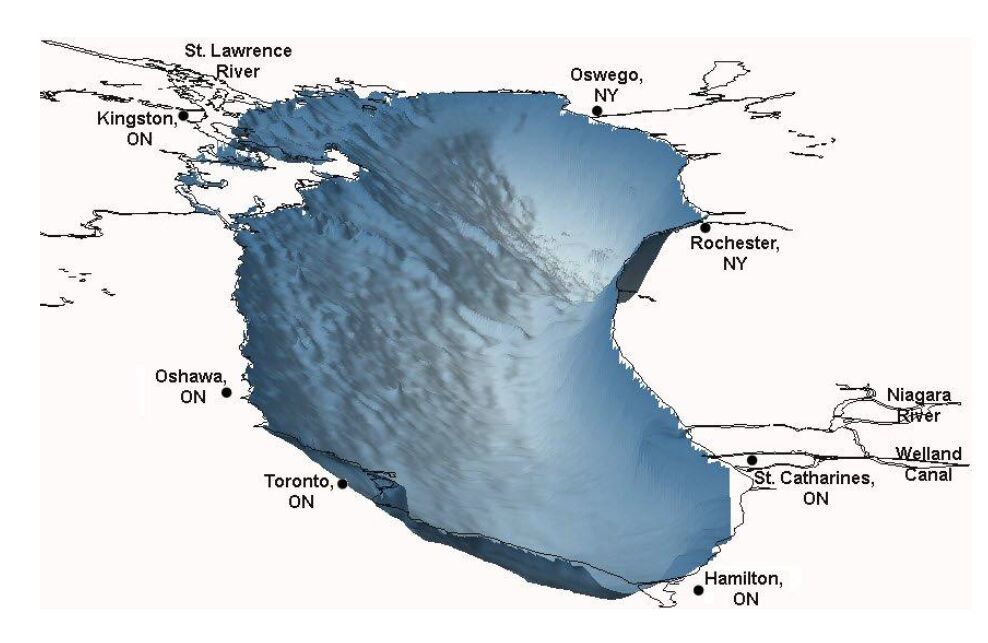
Figures(10) / Tables(6)

K. Wayne Forsythe, Chris H. Marvin, Danielle E. Mitchell, Joseph M. Aversa, Stephen J. Swales, Debbie A. Burniston, James P. Watt, Daniel J. Jakubek, Meghan H. McHenry, Richard R. Shaker. Utilization of bathymetry data to examine lead sediment contamination distributions in Lake Ontario[J]. AIMS Environmental Science, 2016, 3(3): 347-361. doi: 10.3934/environsci.2016.3.347







 DownLoad:
DownLoad: