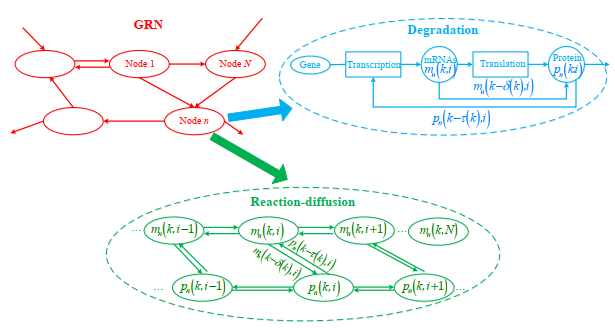
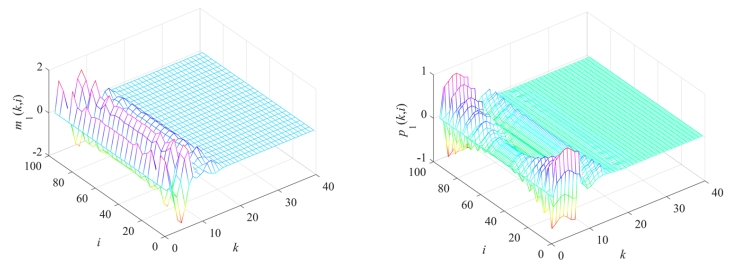
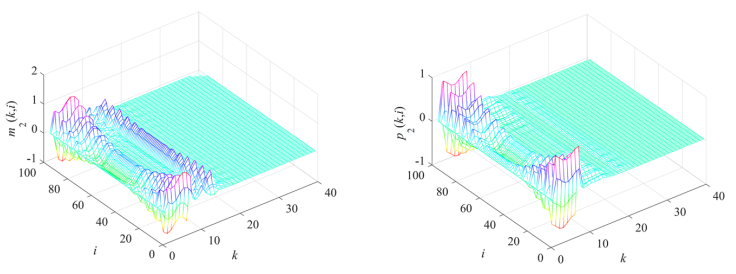
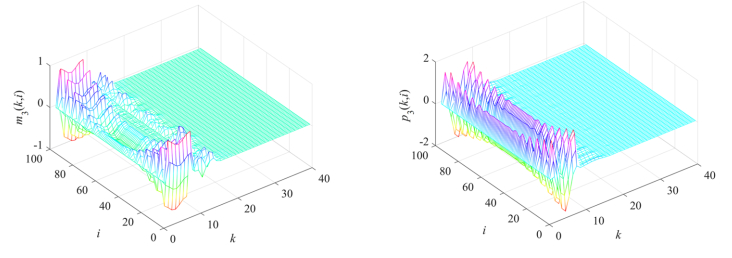
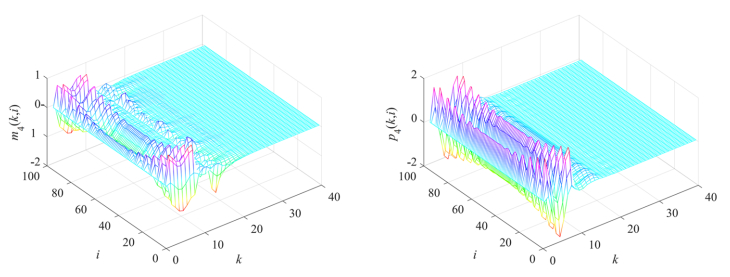
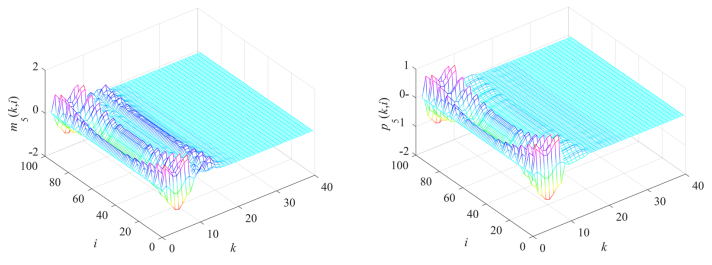
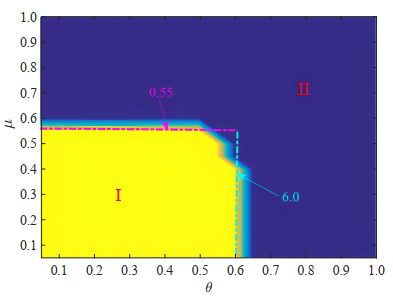
Gene regulatory networks (GRNs) play a crucial role in biological processes, with their dynamic behaviors heavily influenced by the spatial organization of genes. In particular, reaction-diffusion mechanisms govern the coupling between adjacent spatial locations in continuous time GRNs. However, traditional models often ignore the spatial coupling and reaction-diffusion properties of these networks, especially in discrete-time settings. In order to solve this problem, a new discrete-time gene regulatory network model is proposed in this paper, which explicitly considers the mutual coupling between adjacent spatial positions. To ensure the passivity of the proposed model, delay-dependent stability criteria are established by constructing appropriate Lyapunov-Krasovskii functions formulated in terms of linear matrix inequalities. To showcase the effectiveness and validity of this approach, a numerical example is presented in this paper. The results reveal that the model accurately captures the spatial coupling and reaction-diffusion nature of gene regulatory networks in discrete time settings.
Citation: Yongwei Yang, Yang Yu, Chunyun Xu, Chengye Zou. Passivity analysis of discrete-time genetic regulatory networks with reaction-diffusion coupling and delay-dependent stability criteria[J]. Electronic Research Archive, 2025, 33(5): 3111-3134. doi: 10.3934/era.2025136
Gene regulatory networks (GRNs) play a crucial role in biological processes, with their dynamic behaviors heavily influenced by the spatial organization of genes. In particular, reaction-diffusion mechanisms govern the coupling between adjacent spatial locations in continuous time GRNs. However, traditional models often ignore the spatial coupling and reaction-diffusion properties of these networks, especially in discrete-time settings. In order to solve this problem, a new discrete-time gene regulatory network model is proposed in this paper, which explicitly considers the mutual coupling between adjacent spatial positions. To ensure the passivity of the proposed model, delay-dependent stability criteria are established by constructing appropriate Lyapunov-Krasovskii functions formulated in terms of linear matrix inequalities. To showcase the effectiveness and validity of this approach, a numerical example is presented in this paper. The results reveal that the model accurately captures the spatial coupling and reaction-diffusion nature of gene regulatory networks in discrete time settings.

| [1] |
J. Hasty, D. McMillen, F. Isaacs, J. J. Collins, Computational studies of gene regulatory networks: in numero molecular biology, Nat. Rev. Genet., 2 (2001), 268–279. https://doi.org/10.1038/35066056 doi: 10.1038/35066056

|
| [2] |
N. Bernaola, M. Michiels, P. Larrañaga, C. Bielza, Learning massive interpretable gene regulatory networks of the human brain by merging Bayesian networks, PloS Comput. Biol., 19 (2023), 1011443. https://doi.org/10.1371/journal.pcbi.1011443 doi: 10.1371/journal.pcbi.1011443
|
| [3] |
J. Musilova, Z. Vafek, B. L. Puniya, R. Zimmer, T. Helikar, K. Sedlar, Augusta: From RNA-Seq to gene regulatory networks and Boolean models, Comput. Struct. Biotechnol. J., 23 (2024), 783–790. https://doi.org/10.1016/j.csbj.2024.01.013 doi: 10.1016/j.csbj.2024.01.013
|
| [4] |
G. Karlebach, P. N. Robinson, Computing minimal Boolean models of gene regulatory networks, J. Comput. Biol., 31 (2024), 117–127. https://doi.org/10.1089/cmb.2023.0122 doi: 10.1089/cmb.2023.0122
|
| [5] |
I. Hossai, V. Fanfani, J. Fischer, J. Quackenbush, R. Burkholz, Biologically informed NeuralODEs for genome-wide regulatory dynamics, Genome Biol., 25 (2024), 127. https://doi.org/10.1186/s13059-024-03264-0 doi: 10.1186/s13059-024-03264-0
|
| [6] |
X. She, L. Wang, Y. Zhang, Finite-time stability of genetic regulatory networks with nondifferential delays, IEEE Trans. Circuits Syst. II: Express Briefs, 70 (2023), 2107–2111. https://doi.org/10.1109/TCSII.2022.3233797 doi: 10.1109/TCSII.2022.3233797
|
| [7] |
T. Zhang, Y. Li, Exponential Euler scheme of multi-delay Caputo-Fabrizio fractional-order differential equations, Appl. Math. Lett., 124 (2022), 107709. https://doi.org/10.1016/j.aml.2021.107709 doi: 10.1016/j.aml.2021.107709
|
| [8] |
A. Polynikis, S. J. Hogan, M. D. Bernardo, Comparing different ODE modelling approaches for gene regulatory networks, J. Theor. Biol., 261 (2009), 511–530. https://doi.org/10.1016/j.jtbi.2009.07.040 doi: 10.1016/j.jtbi.2009.07.040
|
| [9] |
M. Kchaou, G. Narayanan, M. S. Ali, S. Sanober, G. Rajchakit, B. Priya, Finite-time Mittag-Leffler synchronization of delayed fractional-order discrete-time complex-valued genetic regulatory networks: Decomposition and direct approaches, Inf. Sci., 664 (2024), 120337. https://doi.org/10.1016/j.ins.2024.120337 doi: 10.1016/j.ins.2024.120337
|
| [10] |
K. Xie, C. Zhang, S. Lee, Y. Liu, C. Zhai, Sector bound-dependent matrix-separation-based inequality and its application to stability analysis of discrete-time delayed genetic regulatory networks, IEEE Trans. Circuits Syst. II: Express Briefs, 71 (2024), 3785–3789. https://doi.org/10.1109/TCSII.2024.3367777 doi: 10.1109/TCSII.2024.3367777
|
| [11] |
K. Mathiyalagan, R. Sakthivel, Robust stabilization and H∞ control for discrete-time stochastic genetic regulatory networks with time delays, Can. J. Phys., 90 (2012), 939–953. https://doi.org/10.1139/p2012-088 doi: 10.1139/p2012-088
|
| [12] |
Q. Li, H. Q. Wei, D. Hua, J. Wang, J. Yang, Stabilization of semi-Markovian jumping uncertain complex-valued networks with time-varying delay: A sliding-mode control approach, Neural Process Lett., 56 (2024), 111. https://doi.org/10.1007/s11063-024-11585-1 doi: 10.1007/s11063-024-11585-1
|
| [13] |
G. Narayanan, M. S. Ali, H. Alsulami, T. Saeed, B. Ahmad Synchronization of T–S fuzzy fractional-order discrete-time complex-valued molecular models of mRNA and protein in regulatory mechanisms with leakage effects, Neural Process Lett., 55 (2023), 3305–3331. https://doi.org/10.1007/s11063-022-11010-5 doi: 10.1007/s11063-022-11010-5
|
| [14] |
H. Wei, K. Zhang, M. Zhang, Q. Li, J. Wang, Dissipative synchronization of Semi-Markovian jumping delayed neural networks under random deception attacks: An event-triggered impulsive control strategy, J. Franklin Inst., 361 (2024), 106835. https://doi.org/10.1016/j.jfranklin.2024.106835 doi: 10.1016/j.jfranklin.2024.106835
|
| [15] |
S. Pandiselvi, R. Raja, J. Cao, G. Rajchakit, B. Ahmad, Approximation of state variables for discrete-time stochastic genetic regulatory networks with leakage, distributed, and probabilistic measurement delays: A robust stability problem, Adv. Differ. Equations, 2018 (2018), 123. https://doi.org/10.1186/s13662-018-1569-z doi: 10.1186/s13662-018-1569-z
|
| [16] |
Y. Zhao, H. Wu, Fixed/Prescribed stability criterions of stochastic system with time-delay, AIMS Math., 9 (2024), 14425–14453. https://doi.org/10.3934/math.2024701 doi: 10.3934/math.2024701
|
| [17] |
T. Zhang, Z. Li, Switching clusters' synchronization for discrete space-time complex dynamical networks via boundary feedback controls, Pattern Recognit., 143 (2023), 109763. https://doi.org/10.1016/j.patcog.2023.109763 doi: 10.1016/j.patcog.2023.109763
|
| [18] |
R. Liang, J. Wang, PD control for passivity of coupled reaction-diffusion neural networks with multiple state couplings or spatial diffusion couplings, Neurocomputing, 489 (2022), 558–569. https://doi.org/10.1016/j.neucom.2021.12.070 doi: 10.1016/j.neucom.2021.12.070
|
| [19] |
P. U. Avila, T. Padvitski, A. C. Leote, H. Chen, J. Saez-Rodriguez, M. Kann, et al., Gene regulatory networks in disease and ageing, Nat. Rev. Nephrol., 20 (2024), 616–633. https://doi.org/10.1038/s41581-024-00849-7 doi: 10.1038/s41581-024-00849-7
|
| [20] |
P. Jothiappan, M. Kalidass, Robust passivity analysis of stochastic genetic regulatory networks with Levy noise, Int. J. Control Autom. Syst., 20 (2022), 3241–3251. https://doi.org/10.1007/s12555-021-0552-8 doi: 10.1007/s12555-021-0552-8
|
| [21] |
Y. Qin, F. Li, J. Wang, H. Shen, Extended dissipative synchronization of reaction-diffusion genetic regulatory networks based on sampled-data control, Neural Process Lett., 55 (2023), 3169–3183. https://doi.org/10.1007/s11063-022-11003-4 doi: 10.1007/s11063-022-11003-4
|
| [22] |
X. Song, X. Li, S. Song, C. K. Ahn, State observer design of coupled genetic regulatory networks with Reaction-diffusion terms via time-space sampled-data communications, IEEE/ACM Trans. Comput. Biol. Bioinf., 19 (2022), 3704–3714. https://doi.org/10.1109/TCBB.2021.3114405 doi: 10.1109/TCBB.2021.3114405
|
| [23] |
Y. Zhang, H. Liu, H. Yan, J. Zhou, Oscillatory behaviors in genetic regulatory networks mediated by microRNA with time delays and reaction-diffusion terms, IEEE Trans. Nanobiosci., 16 (2017), 166–176. https://doi.org/10.1109/TNB.2017.2675446 doi: 10.1109/TNB.2017.2675446
|
| [24] |
C. Ma, Q. Zeng, L. Zhang, Y. Zhu, Passivity and passification for Markov jump genetic regulatory networks with time-varying delays, Neurocomputing, 136 (2014), 321–326. https://doi.org/10.1016/j.neucom.2013.12.028 doi: 10.1016/j.neucom.2013.12.028
|
| [25] |
X. Zhang, Y. Han, L. Wu, Y. Wang, State estimation for delayed genetic regulatory networks with reaction-diffusion terms, IEEE Trans. Neural Networks Learn. Syst., 29 (2018), 299–309. https://doi.org/10.1109/TNNLS.2016.2618899 doi: 10.1109/TNNLS.2016.2618899
|
Figures(7) / Tables(1)
Yongwei Yang, Yang Yu, Chunyun Xu, Chengye Zou. Passivity analysis of discrete-time genetic regulatory networks with reaction-diffusion coupling and delay-dependent stability criteria[J]. Electronic Research Archive, 2025, 33(5): 3111-3134. doi: 10.3934/era.2025136







 DownLoad:
DownLoad: