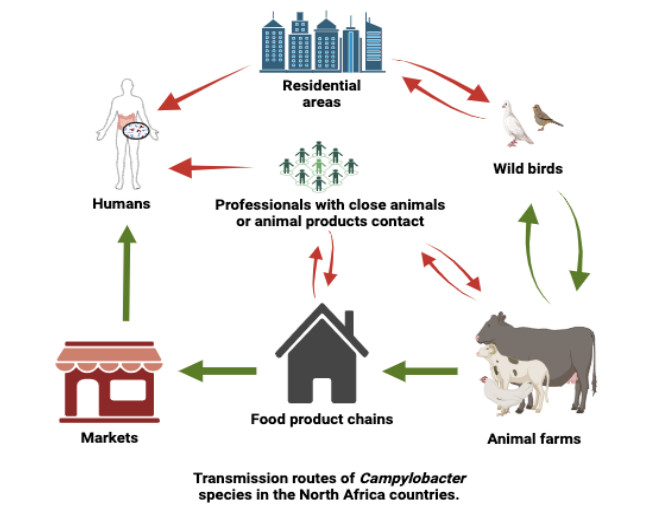
Foodborne bacterial infections, particularly those caused by contaminated food sources, pose significant public health challenges and result in substantial economic losses. This review aims to provide insights into recent literature on the prevalence of Campylobacter spp. in North African food supply chains and their pathogenicity. Additionally, it seeks to summarize the available information on health-related issues and the current state of antibiotic resistance. The reviewed evidence highlights a gap in our understanding of the prevalence of Campylobacter spp. in North African food supplies. Molecular characterization efforts to identify the sources of Campylobacter spp. are limited, and there are few surveys that have specifically targeted this bacterium in the food supply. While qualitative data indicates either the presence or absence of Campylobacter spp., quantitative data on the actual amounts of these bacteria in chicken meat supplies across North African countries are notably lacking. Despite frequent reports of Campylobacter spp. in animal-derived foods, the literature reviewed emphasizes the ongoing challenge that Campylobacter spp. pose to food safety and public health in North Africa.
Citation: Mohamed-Yousif Ibrahim Mohamed. Campylobacteriosis in North Africa[J]. AIMS Agriculture and Food, 2024, 9(3): 801-821. doi: 10.3934/agrfood.2024043
Foodborne bacterial infections, particularly those caused by contaminated food sources, pose significant public health challenges and result in substantial economic losses. This review aims to provide insights into recent literature on the prevalence of Campylobacter spp. in North African food supply chains and their pathogenicity. Additionally, it seeks to summarize the available information on health-related issues and the current state of antibiotic resistance. The reviewed evidence highlights a gap in our understanding of the prevalence of Campylobacter spp. in North African food supplies. Molecular characterization efforts to identify the sources of Campylobacter spp. are limited, and there are few surveys that have specifically targeted this bacterium in the food supply. While qualitative data indicates either the presence or absence of Campylobacter spp., quantitative data on the actual amounts of these bacteria in chicken meat supplies across North African countries are notably lacking. Despite frequent reports of Campylobacter spp. in animal-derived foods, the literature reviewed emphasizes the ongoing challenge that Campylobacter spp. pose to food safety and public health in North Africa.

| [1] |
EFSA (2022) The European Union One Health 2021 Zoonoses Report. EFSA J 20: e07666. https://doi.org/10.2903/j.efsa.2022.7666 doi: 10.2903/j.efsa.2022.7666

|
| [2] |
EFSA, ECDC (2022) The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J 20: e07209. https://doi.org/10.2903/j.efsa.2022.7209 doi: 10.2903/j.efsa.2022.7209
|
| [3] |
EFSA (2019) The European Union One Health 2018 Zoonoses Report. EFSA Journal 17: e05926. https://doi.org/10.2903/j.efsa.2019.5926 doi: 10.2903/j.efsa.2019.5926
|
| [4] |
Tack DM, Ray L, Griffin PM, et al. (2020) Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. Sites, 2016–2019. Morb Mortal Wkly Rep 69: 509–514. https://doi.org/10.15585/mmwr.mm6917a1 doi: 10.15585/mmwr.mm6917a1
|
| [5] | WHO (2015) WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Bur‐ den Epidemiology Reference Group 2007–2015. World Health Organization, Geneva, Switzerland. |
| [6] |
Mohamed M-YI, Habib I (2023) Pathogenic E. coli in the food chain across the Arab countries: A descriptive review. Foods 12: 3726. https://doi.org/10.3390/foods12203726 doi: 10.3390/foods12203726
|
| [7] |
Rafei R, Hawli M, Osman M, et al. (2020) Distribution of EMM types and macrolide resistance determinants among group A Streptococci in the Middle East and North Africa region. J Global Antimicrob Resist 22: 334–348. https://doi.org/10.1016/j.jgar.2020.02.005 doi: 10.1016/j.jgar.2020.02.005
|
| [8] |
Paré G, Trudel MC, Jaana M, et al. (2015) Synthesizing information systems knowledge: A typology of literature reviews. Inf Manage 52: 183–199. https://doi.org/10.1016/j.im.2014.08.008 doi: 10.1016/j.im.2014.08.008
|
| [9] |
Mohamed M-YI, Habib I, Khalifa HO (2024) Salmonella in the food chain within the Gulf Cooperation Council countries. AIMS Microbiol 10: 468–488. https://doi.org/10.3934/microbiol.2024023 doi: 10.3934/microbiol.2024023
|
| [10] |
Butzler JP (2004) Campylobacter, from obscurity to celebrity. Clin Microbiol Infect 10: 868–876. https://doi.org/10.1111/j.1469-0691.2004.00983.x doi: 10.1111/j.1469-0691.2004.00983.x
|
| [11] |
Chon JW, Seo KH, Kim B, et al. (2020) Advanced methods for isolating from and confirming Campylobacter spp. in milk and dairy products: Review. J Dairy Sci Biotechnol 38: 121–133. https://doi.org/10.22424/jdsb.2020.38.3.121 doi: 10.22424/jdsb.2020.38.3.121
|
| [12] |
Parker CT, Cooper KK, Schiaffino F, et al. (2021) Genomic characterization of Campylobacter jejuni adapted to the guinea pig (Cavia porcellus) host. Front Cell Infect Microbiol 11: 607747. https://doi.org/10.3389/fcimb.2021.607747 doi: 10.3389/fcimb.2021.607747
|
| [13] |
Silva MF, Pereira G, Carneiro C, et al. (2020) Campylobacter portucalensis sp. nov., a new species of Campylobacter isolated from the preputial mucosa of bulls. PLoS One 15: e0227500. https://doi.org/10.1371/journal.pone.0227500 doi: 10.1371/journal.pone.0227500
|
| [14] |
Ammar AM, El-Naenaeey E-SY, El-Malt RMS, et al. (2021) Prevalence, antimicrobial susceptibility, virulence and genotyping of Campylobacter jejuni with a special reference to the anti-virulence potential of eugenol and beta-resorcylic acid on some multi-drug resistant isolates in Egypt. Animals 1: 3. https://doi.org/10.3390/ani11010003 doi: 10.3390/ani11010003
|
| [15] | Martora F, Pagliuca C, Della Pepa ME, et al. (2020) Campylobacter jejuni bacteremia in Italian pediatric patients with acute lymphoblastic leukemia: Report of two cases. New Microbiol 43: 1121–7138. |
| [16] |
Zhang XH, Ahmad W, Zhu XY, et al. (2021) Viable but nonculturable bacteria and their resuscitation: implications for cultivating uncultured marine microorganisms. Mar Life Sci Technol 3: 189–203. https://doi.org/10.1007/s42995-020-00041-3 doi: 10.1007/s42995-020-00041-3
|
| [17] |
Baali M, Lounis M, Al Amir HL, et al. (2020) Prevalence, seasonality, and antimicrobial resistance of thermotolerant Campylobacter isolated from broiler farms and slaughterhouses in East Algeria. Vet World 13: 1221–1228. https://doi.org/10.14202/vetworld.2020.1221-1228 doi: 10.14202/vetworld.2020.1221-1228
|
| [18] | Pires SM, Devleesschauwer B (2021) Estimates of global disease burden associated with foodborne pathogens. Foodborne Infections and Intoxications, Elsevier, 3–17. https://doi.org/10.1016/B978-0-12-819519-2.00020-7 |
| [19] |
Senok A, Yousif A, Mazi W, et al. (2007) Pattern of antibiotic susceptibility in Campylobacter jejuni isolates of human and poultry origin. Jpn J Infect Dis 60: 1–4. https://doi.org/10.7883/yoken.JJID.2007.1 doi: 10.7883/yoken.JJID.2007.1
|
| [20] |
Habib I, Mohamed MYI, Khan M (2021) Current state of Salmonella, Campylobacter and listeria in the food chain across the Arab countries: A descriptive review. Foods 10: 2369. https://doi.org/10.3390/foods10102369 doi: 10.3390/foods10102369
|
| [21] | Habib I, Mohamed MYI (2022) Foodborne infections in the Middle East. Food Safety in the Middle East, Elsevier, 71–107. https://doi.org/10.1016/B978-0-12-822417-5.00005-2 |
| [22] |
Ibrahim Mohamed MY (2021) Occurrence of antimicrobial resistance in foodborne bacteria (Campylobacter and E. coli): A food safety issue and public health hazard. Nutri Food Sci Int J 11: 555801. https://doi.org/10.1016/B978-0-12-822417-5.00005-2 doi: 10.1016/B978-0-12-822417-5.00005-2
|
| [23] |
Mohamed M-YI, Abu J, Abdul-Aziz S, et al. (2021) Public health significance of Campylobacter jejuni. J Biosci Med (Irvine) 09: 100–112. https://doi.org/10.4236/jbm.2021.98009 doi: 10.4236/jbm.2021.98009
|
| [24] |
Mohamed M-YI, Abu J, Abdul-Aziz S, et al. (2019) Occurrence of antibiotic resistant C. jejuni and E. coli in wild birds, chickens, environment and humans from Orang Asli Villages in Sungai Siput, Perak, Malaysia. Am J Anim Vet Sci 14: 158–169. https://doi.org/10.3844/ajavsp.2019.158.169 doi: 10.3844/ajavsp.2019.158.169
|
| [25] |
Amjad M, Zia U-U-R (2023) Poultry as A Source and Reservoir for Campylobacteriosis. Eur J Vet Med 3: 11–17. https://doi.org/10.24018/ejvetmed.2023.3.1.87 doi: 10.24018/ejvetmed.2023.3.1.87
|
| [26] |
Habib I, Ibrahim Mohamed MY, Ghazawi A, et al. (2023) Genomic characterization of molecular markers associated with antimicrobial resistance and virulence of the prevalent Campylobacter coli isolated from retail chicken meat in the United Arab Emirates. Curr Res Food Sci 6: 100434. https://doi.org/10.1016/j.crfs.2023.100434 doi: 10.1016/j.crfs.2023.100434
|
| [27] |
Mohamed M-YI, Abu J, Aziz SA, et al. (2022) Occurrence of antibiotic resistant C. jejuni and E. coli in wild birds, chickens, humans, and the environment in Malay villages, Kedah, Malaysia. Vet Med (Praha) 67: 298–308. https://doi.org/10.17221/102/2021-VETMED doi: 10.17221/102/2021-VETMED
|
| [28] |
Colles FM, Ali JS, Sheppard SK, et al. (2011) Campylobacter populations in wild and domesticated Mallard ducks (Anas platyrhynchos). Environ Microbiol Rep 3: 574–580. https://doi.org/10.1111/j.1758-2229.2011.00265.x doi: 10.1111/j.1758-2229.2011.00265.x
|
| [29] |
Cribb DM, Varrone L, Wallace RL, et al. (2022) Risk factors for campylobacteriosis in Australia: Outcomes of a 2018–2019 case-control study. BMC Infect Dis 22: 586. https://doi.org/10.1186/s12879-022-07553-6 doi: 10.1186/s12879-022-07553-6
|
| [30] |
Mohamed M-YI, Abdul-Aziz S, Jalila A, et al. (2019) Occurrence of antibiotic resistant Campylobacter in wild birds and poultry. Malays J Microbiol 15: 143–151. http://dx.doi.org/10.21161/mjm.180096 doi: 10.21161/mjm.180096
|
| [31] |
Gras LM, Smid JH, Wagenaar JA, et al. (2013) Increased risk for Campylobacter jejuni and C. coli infection of pet origin in dog owners and evidence for genetic association between strains causing infection in humans and their pets. Epidemiol Infect 141: 2526–2535. https://doi.org/10.1017/S0950268813000356 doi: 10.1017/S0950268813000356
|
| [32] | Mohamed M-Y-I (2021) Occurrence of Campylobacter jejuni in poultry meats. Nov Res Sci 8: 1–3. |
| [33] |
Benamar I, Nauta M, Cherif-Antar A, et al. (2021) Quantitative risk assessment of Campylobacter in döner kebab consumed in the west of Algeria. Microb Risk Anal 19: 100172. https://doi.org/10.1016/j.mran.2021.100172 doi: 10.1016/j.mran.2021.100172
|
| [34] |
Messad S, Hamdi T-M, Bouhamed R, et al. (2014) Frequency of contamination and antimicrobial resistance of thermotolerant Campylobacter isolated from some broiler farms and slaughterhouses in the region of Algiers. Food Control 40: 324–328. https://doi.org/10.1016/j.foodcont.2013.12.016 doi: 10.1016/j.foodcont.2013.12.016
|
| [35] |
El-Naenaeey E, Abd El-Hamid M, Khalifa E (2021) Prevalence and antibiotic resistanc patterns of Campylobacter species isolated from different sources in Eygpt. J Microbiol, Biotechnol Food Sci 10: e3723. https://doi.org/10.15414/jmbfs.3723 doi: 10.15414/jmbfs.3723
|
| [36] |
Abd El-Tawab AA, Ammar AM, Ahmed HA, et al. (2015) Bacteriological and molecular identification of Campylobacter species in chickens and humans, at Zagazig City, Egypt. Benha Vet Med J 28: 17–26. https://doi.org/10.21608/bvmj.2015.32523 doi: 10.21608/bvmj.2015.32523
|
| [37] |
Zeinhom MMA, Abdel-Latef GK, Corke H (2021) Prevalence, characterization, and control of Campylobacter jejuni isolated from raw milk, cheese, and human stool samples in Beni-Suef governorate, Egypt. Foodborne Pathog Dis 18: 322–330. https://doi.org/10.1089/fpd.2020.2895 doi: 10.1089/fpd.2020.2895
|
| [38] |
Barakat AMA, Abd El-Razik KA, Elfadaly HA, et al. (2020) Prevalence, molecular detection, and virulence gene profiles of Campylobacter species in humans and foods of animal origin. Vet World 13: 1430–1438. https://doi.org/10.14202/vetworld.2020.1430-1438 doi: 10.14202/vetworld.2020.1430-1438
|
| [39] |
El-Zamkan MA, Abdel Hameed KG (2016) Prevalence of Campylobacter jejuni and Campylobacter coli in raw milk and some dairy products. Vet World 9: 1147–1151. https://doi.org/10.14202/vetworld.2016.1147-1151 doi: 10.14202/vetworld.2016.1147-1151
|
| [40] |
El Baaboua A, El Maadoudi M, Bouyahya A, et al. (2021) Prevalence and antimicrobial profiling of Campylobacter spp. isolated from meats, animal, and human feces in Northern of Morocco. Int J Food Microbiol 349: 109202. https://doi.org/10.1016/j.ijfoodmicro.2021.109202 doi: 10.1016/j.ijfoodmicro.2021.109202
|
| [41] | Jouahri M, Asehraou A, Karib H, et al. (2007) Prevalence and control of thermo‐tolerant Campylobacter species in raw poultry meat in Morocco. Meso Prvi Hrvat Časopis Mesu 9: 262–267. |
| [42] |
Béjaoui A, Gharbi M, Bitri S, et al. (2022) Virulence profiling, multidrug resistance and molecular mechanisms of Campylobacter strains from chicken carcasses in Tunisia. Antibiotics 11: 830. https://doi.org/10.3390/antibiotics11070830 doi: 10.3390/antibiotics11070830
|
| [43] |
Gharbi M, Béjaoui A, Ben Hamda C, et al. (2022) Campylobacter spp. in eggs and laying hens in the North-East of Tunisia: High prevalence and multidrug-resistance phenotypes. Vet Sci 9: 108. https://doi.org/10.3390/vetsci9030108 doi: 10.3390/vetsci9030108
|
| [44] |
Jribi H, Sellami H, Mariam S, et al. (2017) Isolation and identification of Campylobacter spp. from poultry and poultry by-products in Tunisia by conventional culture method and multiplex real-time PCR. J Food Prot 80: 1623–1627. https://doi.org/10.4315/0362-028X.JFP-16-321 doi: 10.4315/0362-028X.JFP-16-321
|
| [45] |
Sayed ASM, Ibrahim AI, Sobhy MM, et al. (2023) Circulation of thermophilic Campylobacter in pigeons, turkeys, and humans at live bird markets in Egypt. Front Vet Sci 10: 1150077. https://doi.org/10.3389/fvets.2023.1150077 doi: 10.3389/fvets.2023.1150077
|
| [46] |
Khalil MR, Amira AM, Kafafy MH, et al. (2020) Molecular characterization of Campylobacter species from Turkeys flocks in delta governments. Assiut Vet Med J 66: 111–117. https://doi.org/10.21608/avmj.2020.167255 doi: 10.21608/avmj.2020.167255
|
| [47] |
Lobna MAS, Nashwa OKh, Barakat AMA, et al. (2019) Antimicrobial resistance of Campylobacter jejuni isolated from chicken, some animal products and human in Kalyoubia, Egypt, with special reference to its viability. Benha Vet Med J 36: 156–163. https://doi.org/10.21608/bvmj.2019.103407 doi: 10.21608/bvmj.2019.103407
|
| [48] |
Abushahba MF, Ahmed SO, Ibrahim AA, et al. (2018) Prevalence of zoonotic species of Campylobacter in broiler chicken and humans in Assiut governorate, Egypt. Approaches Poult Dairy Vet Sci 3: 260–268. https://doi.org/10.31031/APDV.2018.03.000568 doi: 10.31031/APDV.2018.03.000568
|
| [49] | Abd El-Aziz DM, Abd-Allah SMS (2016) Incidence of Campylobacter species in wholesale chicken carcasses and chicken meat products in Assiut city, Egypt. Int Food Res J 24: 2660–2665. |
| [50] |
El-Sharoud WM (2009) Prevalence and survival of Campylobacter in Egyptian dairy products. Food Research International 42: 622–626. https://doi.org/10.1016/j.foodres.2009.01.009 doi: 10.1016/j.foodres.2009.01.009
|
| [51] |
Es-Soucratti K, Hammoumi A, Bouchrif B, et al. (2020) Occurrence and antimicrobial resistance of Campylobacter jejuni isolates from poultry in Casablanca-Settat, Morocco. Ital J Food Saf 9: 54–59. https://doi.org/10.4081/ijfs.2020.8692 doi: 10.4081/ijfs.2020.8692
|
| [52] |
Asmai R, Karraouan B, Es-Soucratti K, et al. (2020) Prevalence and antibiotic resistance of Campylobacter coli isolated from broiler farms in the Marrakesh Safi region, Morocco. Vet World 13: 1892–1897. https://doi.org/10.14202/vetworld.2020.1892-1897 doi: 10.14202/vetworld.2020.1892-1897
|
| [53] |
Gharbi M, Béjaoui A, Ben Hamda C, et al. (2018) Prevalence and antibiotic resistance patterns of Campylobacter spp. isolated from broiler chickens in the North of Tunisia. Biomed Res Int 2018: 1–7. https://doi.org/10.1155/2018/7943786 doi: 10.1155/2018/7943786
|
| [54] |
Brückner V, Fiebiger U, Ignatius R, et al. (2020) Characterization of Arcobacter strains isolated from human stool samples: Results from the prospective German prevalence study Arcopath. Gut Pathog 12: 3. https://doi.org/10.1186/s13099-019-0344-3 doi: 10.1186/s13099-019-0344-3
|
| [55] |
Mihevc M, Koren Krajnc M, Bombek Ihan M, et al. (2021) Helicobacter canis bacteraemia in a rheumatoid arthritis patient treated with tofacitinib: Case report and literature review. Ann Clin Microbiol Antimicrob 20: 22. https://doi.org/10.1186/s12941-021-00426-x doi: 10.1186/s12941-021-00426-x
|
| [56] |
Eryıldız C, Şakru N, Tabakçıoğlu K, et al. (2022) Molecular identification of Campylobacter species isolated from patients with gastroenteritis in Edirne, Turkey. Cyprus J Med Sci 7: 623–627. https://doi.org/10.4274/cjms.2022.2021-164 doi: 10.4274/cjms.2022.2021-164
|
| [57] |
Ghoneim NH, Abdel-Moein KA-A, Barakat AMAK, et al. (2021) Isolation and molecular characterization of Campylobacter jejuni from chicken and human stool samples in Egypt. Food Sci Technol 41: 195–202. https://doi.org/10.1590/fst.01620 doi: 10.1590/fst.01620
|
| [58] | Abdelhady HAS, Hussein AAA, Amen O, et al. (2024) Genetic diversity, virulence profile of Campylobacter coli and Campylobacter jejuni isolated from poultry and human in Assiut governorate, Egypt. J Adv Vet Res 14: 193–197. |
| [59] |
Lopes GV, Ramires T, Kleinubing NR, et al. (2021) Virulence factors of foodborne pathogen Campylobacter jejuni. Microb Pathog 161: 105265. https://doi.org/10.1016/j.micpath.2021.105265 doi: 10.1016/j.micpath.2021.105265
|
| [60] |
Talukdar PK, Negretti NM, Turner KL, et al. (2020) Molecular dissection of the Campylobacter jejuni CadF and FlpA virulence proteins in binding to host cell fibronectin. Microorganisms 8: 389. https://doi.org/10.3390/microorganisms8030389 doi: 10.3390/microorganisms8030389
|
| [61] |
Jakee JE, Ata NS, Hakim AS, et al. (2015) Prevalence of virulence genes and antimicrobial resistance patterns of Campylobacter species isolated from chicken in Egypt. Asian J Poult Sci 9: 250–261. https://doi.org/10.3923/ajpsaj.2015.250.261 doi: 10.3923/ajpsaj.2015.250.261
|
| [62] |
Gharbi M, Béjaoui A, Ben Hamda C, et al. (2022) Distribution of virulence and antibiotic resistance genes in Campylobacter jejuni and Campylobacter coli isolated from broiler chickens in Tunisia. J Microbiol, Immunol Infect 55: 1273–1282. https://doi.org/10.1016/j.jmii.2021.07.001 doi: 10.1016/j.jmii.2021.07.001
|
| [63] |
Nahar N, Rashid R Bin (2018) Genotypic analysis of the virulence and antibiotic resistance genes in Campylobacter species in silico. J Bioanal Biomed 10: 13–23. https://doi.org/10.4172/1948-593X.1000199 doi: 10.4172/1948-593X.1000199
|
| [64] |
Eryildiz C, Tabakcioglu K, Kuyucuklu G, et al. (2020) Investigation of antimicrobial resistance and virulence genes of Campylobacter isolates from patients in a tertiary hospital in Edirne, Turkey. Indian J Med Microbiol 38: 157–161. https://doi.org/10.4103/ijmm.IJMM_20_78 doi: 10.4103/ijmm.IJMM_20_78
|
| [65] | Tegtmeyer N, Sharafutdinov I, Harrer A, et al. (2021) Campylobacter virulence factors and molecular host-pathogen interactions, Fighting Campylobacter Infections. Current Topics in Microbiology and Immunology, 169–202. https://doi.org/10.1007/978-3-030-65481-8_7 |
| [66] |
Alam A, Bröms JE, Kumar R, et al. (2021) The role of ClpB in bacterial stress responses and virulence. Front Mol Biosci 8: 668910. https://doi.org/10.3389/fmolb.2021.668910 doi: 10.3389/fmolb.2021.668910
|
| [67] |
Koolman L, Whyte P, Burgess C, et al. (2015) Distribution of virulence-associated genes in a selection of Campylobacter isolates. Foodborne Pathog Dis 12: 424–432. https://doi.org/10.1089/fpd.2014.1883 doi: 10.1089/fpd.2014.1883
|
| [68] |
Saif NA, Cobo‐Díaz JF, Elserafy M, et al. (2022) A pilot study revealing host‐associated genetic signatures for source attribution of sporadic Campylobacter jejuni infection in Egypt. Transbound Emerg Dis 69: 1847–1861. https://doi.org/10.1111/tbed.14165 doi: 10.1111/tbed.14165
|
| [69] |
Gharbi M, Kamoun S, Hkimi C, et al. (2022) Relationships between virulence genes and antibiotic resistance phenotypes/genotypes in Campylobacter spp. isolated from layer hens and eggs in the North of Tunisia: Statistical and computational insights. Foods 11: 3554. https://doi.org/10.3390/foods11223554 doi: 10.3390/foods11223554
|
| [70] |
Jribi H, Sellami H, Hassena A Ben, et al. (2017) Prevalence of putative virulence genes in Campylobacter and Arcobacter species isolated from poultry and poultry by-products in Tunisia. J Food Prot 80: 1705–1710. https://doi.org/10.4315/0362-028X.JFP-16-509 doi: 10.4315/0362-028X.JFP-16-509
|
| [71] |
Dias TS, Panzenhagen P, Figueira ADA, et al. (2023) Genomic characterisation of Campylobacter jejuni Cj26: A high-level ciprofloxacin/erythromycin-resistant strain isolated from a poultry carcass in southern Brazil. J Glob Antimicrob Resist 34: 1–4. https://doi.org/10.1016/j.jgar.2023.05.009 doi: 10.1016/j.jgar.2023.05.009
|
| [72] |
Gao F, Tu L, Chen M, et al. (2023) Erythromycin resistance of clinical Campylobacter jejuni and Campylobacter coli in Shanghai, China. Front Microbiol 14: 1145581. https://doi.org/10.1016/j.jgar.2023.05.009 doi: 10.1016/j.jgar.2023.05.009
|
| [73] |
Mohamed M-YI, Abu J, Zakaria Z, et al. (2022) Multi-drug resistant pathogenic Escherichia coli isolated from wild birds, chicken, and the environment in Malaysia. Antibiotics 11: 1275. https://doi.org/10.3390/antibiotics11101275 doi: 10.3390/antibiotics11101275
|
Figures(1) / Tables(3)

Mohamed-Yousif Ibrahim Mohamed. Campylobacteriosis in North Africa[J]. AIMS Agriculture and Food, 2024, 9(3): 801-821. doi: 10.3934/agrfood.2024043







 DownLoad:
DownLoad: