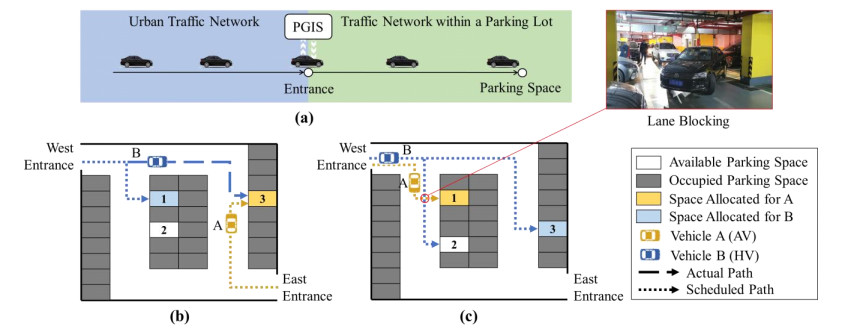
The advent of autonomous vehicles (AVs) poses challenges to parking guidance in mixed driving scenarios involving human-driven vehicles (HVs) and AVs. This study introduced a dynamic and coordinated strategy (DCS) to optimize parking space allocation and path guidance within a mixed driving parking lot, aiming to enhance parking-cruising efficiency. DCS considers the distinctive characteristics of HVs and AVs and dynamically formulates parking guiding schemes based on real-time conditions. The strategy encompasses four main steps: Triggering scheme formulation, identifying preoccupied parking spaces, updating the parking lot traffic network and optimizing the vehicle-path-space matching scheme. A programming model was established to minimize the total remaining cruising time, and iterative optimization was conducted with vehicle loading test based on timing. To elevate computational efficiency, the concept of parking-cruising path tree (PCPT) and its updating method were introduced based on the dynamic shortest path tree algorithm. Comparative analysis of cases and simulations demonstrated the efficacy of DCS in mitigating parking-cruising duration of different types of vehicles and minimizing forced delays arising from lane blocking. Notably, the optimization effect is particularly significant for vehicles with extended cruising durations or in parking lots with low AV penetration rates and high saturation, with an achievable optimization rate reaching up to 18%. This study addressed challenges related to drivers' noncompliance with guidance and lane blocking, thereby improving overall operational efficiency in mixed driving parking lots.
Citation: Zhiyuan Wang, Chu Zhang, Shaopei Xue, Yinjie Luo, Jun Chen, Wei Wang, Xingchen Yan. Dynamic coordinated strategy for parking guidance in a mixed driving parking lot involving human-driven and autonomous vehicles[J]. Electronic Research Archive, 2024, 32(1): 523-550. doi: 10.3934/era.2024026
The advent of autonomous vehicles (AVs) poses challenges to parking guidance in mixed driving scenarios involving human-driven vehicles (HVs) and AVs. This study introduced a dynamic and coordinated strategy (DCS) to optimize parking space allocation and path guidance within a mixed driving parking lot, aiming to enhance parking-cruising efficiency. DCS considers the distinctive characteristics of HVs and AVs and dynamically formulates parking guiding schemes based on real-time conditions. The strategy encompasses four main steps: Triggering scheme formulation, identifying preoccupied parking spaces, updating the parking lot traffic network and optimizing the vehicle-path-space matching scheme. A programming model was established to minimize the total remaining cruising time, and iterative optimization was conducted with vehicle loading test based on timing. To elevate computational efficiency, the concept of parking-cruising path tree (PCPT) and its updating method were introduced based on the dynamic shortest path tree algorithm. Comparative analysis of cases and simulations demonstrated the efficacy of DCS in mitigating parking-cruising duration of different types of vehicles and minimizing forced delays arising from lane blocking. Notably, the optimization effect is particularly significant for vehicles with extended cruising durations or in parking lots with low AV penetration rates and high saturation, with an achievable optimization rate reaching up to 18%. This study addressed challenges related to drivers' noncompliance with guidance and lane blocking, thereby improving overall operational efficiency in mixed driving parking lots.

| [1] | R. C. Hampshire, D. Shoup, What Share of Traffic is Cruising for Parking?, J. Transp. Econ. Policy, 52 (2018), 184–201. |
| [2] |
G. D. Chiara, A. Goodchild, Do commercial vehicles cruise for parking? Empirical evidence from Seattle, Transp. Policy, 97 (2020), 26–36. https://doi.org/10.1016/j.tranpol.2020.06.013 doi: 10.1016/j.tranpol.2020.06.013

|
| [3] |
H. M. Qin, X. H. Yang, Y. J. Wu, H. Z. Guan, P. F. Wang, N. Shahinpoor, Analysis of parking cruising behaviour and parking location choice, Transp. Plan. Technol., 43 (2020), 717–734. https://doi.org/10.1080/03081060.2020.1805545 doi: 10.1080/03081060.2020.1805545
|
| [4] |
R. Grbić, B. Koch, Automatic vision-based parking slot detection and occupancy classification, Expert Syst. Appl., 225 (2023), 120147. https://doi.org/10.1016/j.eswa.2023.120147 doi: 10.1016/j.eswa.2023.120147
|
| [5] |
J. Parmar, P. Das, S. M. Dave, Study on demand and characteristics of parking system in urban areas: A review, J. Traffic Transp. Eng., 7 (2020), 111–124. https://doi.org/10.1016/j.jtte.2019.09.003 doi: 10.1016/j.jtte.2019.09.003
|
| [6] |
F. Al-Turjman, A. Malekloo, Smart parking in IoT-enabled cities: A survey, Sust. Cities Soc., 49 (2019), 101608. https://doi.org/10.1016/j.scs.2019.101608 doi: 10.1016/j.scs.2019.101608
|
| [7] |
T. Lin, H. Rivano, F. Le Mouël, A survey of smart parking solutions, IEEE Trans. Intell. Transp. Syst., 18 (2017), 3229–3253. https://doi.org/10.1109/TITS.2017.2685143 doi: 10.1109/TITS.2017.2685143
|
| [8] |
X. Di, R. Shi, A survey on autonomous vehicle control in the era of mixed-autonomy: From physics-based to AI-guided driving policy learning, Transp. Res. Pt. C-Emerg. Technol., 125 (2021), 103008. https://doi.org/10.1016/j.trc.2021.103008 doi: 10.1016/j.trc.2021.103008
|
| [9] |
Z. Wang, Y. Shi, W. Tong, Z. Gu, Q. Cheng, Car-following models for human-driven vehicles and autonomous vehicles: A systematic review, J. Transp. Eng., Part A: Syst., 149 (2023), 04023075. https://doi.org/10.1061/JTEPBS.TEENG-7836 doi: 10.1061/JTEPBS.TEENG-7836
|
| [10] | Y. Geng, C. G. Cassandras, Dynamic resource allocation in urban settings: A "smart parking" approach, in 2011 IEEE International Symposium on Computer-Aided Control System Design (CACSD), IEEE, (2011), 1–6. https://doi.org/10.1109/CACSD.2011.6044566 |
| [11] |
T. K. Chan, C. S. Chin, Review of autonomous intelligent vehicles for urban driving and parking, Electronics, 10 (2021), https://doi.org/10.3390/electronics10091021 doi: 10.3390/electronics10091021
|
| [12] |
P. S. Perumal, M. Sujasree, S. Chavhan, D. Gupta, V. Mukthineni, S. R. Shimgekar, et al., An insight into crash avoidance and overtaking advice systems for Autonomous Vehicles: A review, challenges and solutions, Eng. Appl. Artif. Intell., 104 (2021), 104406. https://doi.org/10.1016/j.engappai.2021.104406 doi: 10.1016/j.engappai.2021.104406
|
| [13] |
K. Huang, C. Jiang, P. Li, A. Shan, J. Wan, W. Qin, A systematic framework for urban smart transportation towards traffic management and parking, Electron. Res. Arch., 30 (2022), 4191–4208. https://doi.org/10.3934/era.2022212 doi: 10.3934/era.2022212
|
| [14] |
X. Zhang, C. Zhao, F. Liao, X. Li, Y. Du, Online parking assignment in an environment of partially connected vehicles: A multi-agent deep reinforcement learning approach, Transp. Res. Part C: Emerg. Technol., 138 (2022), 103624. https://doi.org/10.1016/j.trc.2022.103624 doi: 10.1016/j.trc.2022.103624
|
| [15] | K. Ata, A. C. Soh, A. Ishak, H. Jaafar, N. Khairuddin, Smart indoor parking system based on dijkstra's algorithm, Int. J. Integr. Eng, 2 (2019), 13–20. |
| [16] |
L. Yu, H. Jiang, L. Hua, Anti-congestion route planning scheme based on Dijkstra algorithm for automatic valet parking system, Appl. Sci.-Basel, 9 (2019), 5016. https://doi.org/10.3390/app9235016 doi: 10.3390/app9235016
|
| [17] |
M. Wu, H. Jiang, C. A. Tan, Automated parking space allocation during transition with both human-operated and autonomous vehicles, Appl. Sci., 11 (2021), 855. https://doi.org/10.3390/app11020855 doi: 10.3390/app11020855
|
| [18] |
C. Lu, C. Liu, Ecological control strategy for cooperative autonomous vehicle in mixed traffic considering linear stability, J. Intell. Connected Veh., 4 (2021), 115–124. https://doi.org/10.1108/JICV-08-2021-0012 doi: 10.1108/JICV-08-2021-0012
|
| [19] |
E. F. Ozioko, J. Kunkel, F. Stahl, Road intersection coordination scheme for mixed traffic (human-driven and driverless vehicles): A systematic review, J. Adv. Transp., 2022 (2022), 2951999. https://doi.org/10.1155/2022/2951999 doi: 10.1155/2022/2951999
|
| [20] |
A. Schieben, M. Wilbrink, C. Kettwich, R. Madigan, T. Louw, N. Merat, Designing the interaction of automated vehicles with other traffic participants: design considerations based on human needs and expectations, Cogn. Technol. Work, 21 (2019), 69–85. https://doi.org/10.1007/s10111-018-0521-z doi: 10.1007/s10111-018-0521-z
|
| [21] |
H. Shi, D. Chen, N. Zheng, X. Wang, Y. Zhou, B. Ran, A deep reinforcement learning based distributed control strategy for connected automated vehicles in mixed traffic platoon, Transp. Res. Part C: Emerg. Technol., 148 (2023), 104019. https://doi.org/10.1016/j.trc.2023.104019 doi: 10.1016/j.trc.2023.104019
|
| [22] |
S. Gong, L. Du, Cooperative platoon control for a mixed traffic flow including human drive vehicles and connected and autonomous vehicles, Transp. Res. Part B: Methodol., 116 (2018), 25–61. https://doi.org/10.1016/j.trb.2018.07.005 doi: 10.1016/j.trb.2018.07.005
|
| [23] |
Y. He, Y. Liu, L. Yang, X. Qu, Deep adaptive control: Deep reinforcement learning-based adaptive vehicle trajectory control algorithms for different risk levels, IEEE Trans. Intell. Veh., (2023), 1–12. https://doi.org/10.1109/TIV.2023.3303408 doi: 10.1109/TIV.2023.3303408
|
| [24] |
S. Yan, T. Welschehold, D. Büscher, W. Burgard, Courteous behavior of automated vehicles at unsignalized intersections via rreinforcement learning, IEEE Robot. Autom. Lett., 7 (2022), 191–198. https://doi.org/10.1109/LRA.2021.3121807 doi: 10.1109/LRA.2021.3121807
|
| [25] |
S. Aoki, R. Rajkumar, Safe intersection management with cooperative perception for mixed traffic of human-driven and autonomous vehicles, IEEE Open J. Veh. Technol., 3 (2022), 251–265. https://doi.org/10.1109/OJVT.2022.3177437 doi: 10.1109/OJVT.2022.3177437
|
| [26] |
Z. H. Khattak, B. L. Smith, M. D. Fontaine, J. Ma, A. J. Khattak, Active lane management and control using connected and automated vehicles in a mixed traffic environment, Transp. Res. Part C: Emerg. Technol., 139 (2022), 103648. https://doi.org/10.1016/j.trc.2022.103648 doi: 10.1016/j.trc.2022.103648
|
| [27] |
J. Wang, L. Lu, S. Peeta, Z. He, Optimal toll design problems under mixed traffic flow of human-driven vehicles and connected and autonomous vehicles, Transp. Res. Part C: Emerg. Technol., 125 (2021), 102952. https://doi.org/10.1016/j.trc.2020.102952 doi: 10.1016/j.trc.2020.102952
|
| [28] |
W. Yue, C. Li, S. Wang, N. Xue, J. Wu, Cooperative incident management in mixed traffic of CAVs and human-driven vehicles, IEEE Trans. Intell. Transp. Syst., (2023), 1–15. https://doi.org/10.1109/TITS.2023.3289983 doi: 10.1109/TITS.2023.3289983
|
| [29] |
S. Bahrami, M. J. Roorda, Optimal traffic management policies for mixed human and automated traffic flows, Transp. Res. Part A: Policy Pract., 135 (2020), 130–143. https://doi.org/10.1016/j.tra.2020.03.007 doi: 10.1016/j.tra.2020.03.007
|
| [30] |
Z. Guo, D. Z. W. Wang, D. Wang, Managing mixed traffic with autonomous vehicles–A day-to-day routing allocation scheme, Transp. Res. Part C: Emerg. Technol., 140 (2022), 103726. https://doi.org/10.1016/j.trc.2022.103726 doi: 10.1016/j.trc.2022.103726
|
| [31] |
T. Roughgarden, É. Tardos, How bad is selfish routing?, J. ACM, 49 (2002), 236–259. https://doi.org/10.1145/506147.506153 doi: 10.1145/506147.506153
|
| [32] |
L. Ma, S. Qu, L. Song, J. Zhang, J. Ren, Human-like car-following modeling based on online driving style recognition, Electron. Res. Arch., 31 (2023), 3264–3290. https://doi.org/10.3934/era.2023165 doi: 10.3934/era.2023165
|
| [33] |
I. Klein, N. Levy, E. Ben-Elia, An agent-based model of the emergence of cooperation and a fair and stable system optimum using ATIS on a simple road network, Transp. Res. Part C: Emerg. Technol., 86 (2018), 183–201. https://doi.org/10.1016/j.trc.2017.11.007 doi: 10.1016/j.trc.2017.11.007
|
| [34] |
Y. Yang, J. Chen, J. Ye, J. Chen, Y. Luo, Joint optimization of facility layout and spatially differential parking pricing for parking lots, Transp. Res. Record, 2677 (2023), 241–257. https://doi.org/10.1177/03611981221145139 doi: 10.1177/03611981221145139
|
| [35] | L. Cheng, C. Liu, B. Yan, Improved hierarchical A-star algorithm for optimal parking path planning of the large parking lot, in 2014 IEEE International Conference on Information and Automation (ICIA), IEEE, (2014), 695–698. https://doi.org/10.1109/ICInfA.2014.6932742 |
| [36] | X. Wei, Z. Lei, Q. Ze-hua, Z. Yong, Z. Tie-nan, Design of parking lot path guidance system based on multi-objective Point A* Algorithm, Comput. Modernizat., (2020), 40. |
| [37] | W. Wang, M. Wang, Y. Yue, Y. Yang, A multi-objective parking space decision-making and path planning method, in 2021 IEEE International Conference on Unmanned Systems (ICUS), (2021), 672–677. https://doi.org/10.1109/ICUS52573.2021.9641394 |
| [38] | G. Wang, T. Nakanishi, A. Fukuda, Time-varying shortest path algorithm with transit time tuning for parking lot navigation, in TENCON 2015-2015 IEEE Region 10 Conference, IEEE, (2015), 1–6. https://doi.org/10.1109/TENCON.2015.7373015 |
| [39] |
S. Gai, X. Zeng, X. Yue, Z. Yuan, Parking guidance model based on user and system bi⁃level optimization algorithm, J. Jilin Univ. (Eng. Technol. Edit.), 52 (2022), 1344–1352. https://doi.org/10.13229/j.cnki.jdxbgxb20210093 doi: 10.13229/j.cnki.jdxbgxb20210093
|
| [40] |
Y. Chen, T. Wang, X. Yan, C. Wang, An ensemble optimization strategy for dynamic parking-space allocation, IEEE Intell. Transp. Syst. Mag., 15 (2023), 347–362. https://doi.org/10.1109/MITS.2022.3163506 doi: 10.1109/MITS.2022.3163506
|
| [41] |
D. C. Tseng, T. W. Chao, J. W. Chang, Image-based parking guiding using ackermann steering geometry, Appl. Mech. Mater., 437 (2013), 823–826. https://doi.org/10.4028/www.scientific.net/AMM.437.823 doi: 10.4028/www.scientific.net/AMM.437.823
|
| [42] | C. Löper, C. Brunken, G. Thomaidis, S. Lapoehn, P. P. Fouopi, H. Mosebach, et al., Automated valet parking as part of an integrated travel assistance, in 16th International IEEE Conference on Intelligent Transportation Systems (ITSC 2013), (2013), 2341–2348. https://doi.org/10.1109/ITSC.2013.6728577 |
| [43] |
W. Fu, Q. Li, Z. Fan, Research on automatic parking path optimization based on Ackerman model, Acad. J. Comput. Inf. Sci., 5 (2022), 50–57. https://doi.org/10.25236/ajcis.2022.050808 doi: 10.25236/ajcis.2022.050808
|
| [44] | Y. Yao, Z. Feng, Z. An, Automatic parking problem based on path planning, in 2023 IEEE International Conference on Control, Electronics and Computer Technology (ICCECT), (2023), 110–115. https://doi.org/10.1109/ICCECT57938.2023.10141401 |
| [45] | L. Yuan, R. Huang, L. Han, M. Zhou, A Parking Guidance Algorithm Based on Time-Optimal Dynamic Sorting for Underground Parking, in 2018 26th International Conference on Geoinformatics, (2018), 1–6. https://doi.org/10.1109/GEOINFORMATICS.2018.8557079 |
| [46] |
M. Kneissl, A. K. Madhusudhanan, A. Molin, H. Esen, S. Hirche, A multi-vehicle control framework with application to automated valet parking, IEEE Trans. Intell. Transp. Syst., 22 (2021), 5697–5707. https://doi.org/10.1109/TITS.2020.2990294 doi: 10.1109/TITS.2020.2990294
|
| [47] | P. Schörner, M. Conzelmann, T. Fleck, M. Zofka, J. M. Zöllner, Park my car! automated valet parking with different vehicle automation levels by v2x connected smart infrastructure, in 2021 IEEE International Intelligent Transportation Systems Conference (ITSC), (2021), 836–843. https://doi.org/10.1109/ITSC48978.2021.9565095 |
| [48] |
B. Xiao, H. N. Cao, Z. L. Shao, E. H. M. Sha, An efficient algorithm for dynamic shortest path tree update in network routing, J. Commun. Networking, 9 (2007), 499–510. https://doi.org/10.1109/JCN.2007.6182886 doi: 10.1109/JCN.2007.6182886
|
| [49] |
P. Narváez, K. Siu, H. Tzeng, New dynamic SPT algorithm based on a ball-and-string model, IEEE-ACM Trans. Networking, 9 (2001), 706–718. https://doi.org/10.1109/90.974525 doi: 10.1109/90.974525
|
| [50] | Driving Test Contents and Methods, China: Ministry of Public Security of the People's Republic of China. |
| [51] | R. RSEV, Self-Parking cars Tesla v Audi v Ford v BMW - test and comparison-are they any good?, 2022. Available from: https://www.youtube.com/watch?v = nsb2XBAIWyA. |
| [52] | R. RSEV, Self-Parking Tesla Vision v Hyundai Ioniq 5 with remote parking v BMW i3, 2022. Available from: https://www.youtube.com/watch?v = -nBnZ7GqPiM. |
| [53] |
Y. Liu, F. Wu, Z. Liu, K. Wang, F. Wang, X. Qu, Can language models be used for real-world urban-delivery route optimization?, Innovation, 4 (2023), 100520. https://doi.org/10.1016/j.xinn.2023.100520 doi: 10.1016/j.xinn.2023.100520
|
| [54] |
X. Qu, H. Lin, Y. Liu, Envisioning the future of transportation: Inspiration of ChatGPT and large models, Commun. Transp. Res., 3 (2023), 100103. https://doi.org/10.1016/j.commtr.2023.100103 doi: 10.1016/j.commtr.2023.100103
|
| [55] |
H. Lin, Y. Liu, S. Li, X. Qu, How generative adversarial networks promote the development of intelligent transportation systems: A survey, IEEE/CAA J. Autom. Sin., 10 (2023), 1781–1796. https://doi.org/10.1109/JAS.2023.123744 doi: 10.1109/JAS.2023.123744
|
| [56] |
E. Singh, N. Pillay, A study of ant-based pheromone spaces for generation constructive hyper-heuristics, Swarm Evol. Comput., 72 (2022), 101095. https://doi.org/10.1016/j.swevo.2022.101095 doi: 10.1016/j.swevo.2022.101095
|
| [57] |
M. Chen, Y. Tan, SF-FWA: A Self-Adaptive Fast Fireworks Algorithm for effective large-scale optimization, Swarm Evol. Comput., 80 (2023), 101314. https://doi.org/10.1016/j.swevo.2023.101314 doi: 10.1016/j.swevo.2023.101314
|
| [58] |
M. A. Dulebenets, An Adaptive Polyploid Memetic Algorithm for scheduling trucks at a cross-docking terminal, Inf. Sci., 565 (2021), 390–421. https://doi.org/10.1016/j.ins.2021.02.039 doi: 10.1016/j.ins.2021.02.039
|
| [59] |
J. Pasha, A. L. Nwodu, A. M. Fathollahi-Fard, G. Tian, Z. Li, H. Wang, et al., Exact and metaheuristic algorithms for the vehicle routing problem with a factory-in-a-box in multi-objective settings, Adv. Eng. Inf., 52 (2022), 101623. https://doi.org/10.1016/j.aei.2022.101623 doi: 10.1016/j.aei.2022.101623
|
| [60] |
P. Singh, J. Pasha, R. Moses, J. Sobanjo, E. E. Ozguven, M. A. Dulebenets, Development of exact and heuristic optimization methods for safety improvement projects at level crossings under conflicting objectives, Reliab. Eng. Syst. Saf., 220 (2022), 108296. https://doi.org/10.1016/j.ress.2021.108296 doi: 10.1016/j.ress.2021.108296
|
| [61] |
Y. Liu, R. Jia, J. Ye, X. Qu, How machine learning informs ride-hailing services: A survey, Commun. Transp. Res., 2 (2022), 100075. https://doi.org/10.1016/j.commtr.2022.100075 doi: 10.1016/j.commtr.2022.100075
|
| [62] |
M. A. Dulebenets, A Diffused Memetic Optimizer for reactive berth allocation and scheduling at marine container terminals in response to disruptions, Swarm Evol. Comput., 80 (2023), 101334. https://doi.org/10.1016/j.swevo.2023.101334 doi: 10.1016/j.swevo.2023.101334
|










Figures(11) / Tables(4)
Zhiyuan Wang, Chu Zhang, Shaopei Xue, Yinjie Luo, Jun Chen, Wei Wang, Xingchen Yan. Dynamic coordinated strategy for parking guidance in a mixed driving parking lot involving human-driven and autonomous vehicles[J]. Electronic Research Archive, 2024, 32(1): 523-550. doi: 10.3934/era.2024026







 DownLoad:
DownLoad: