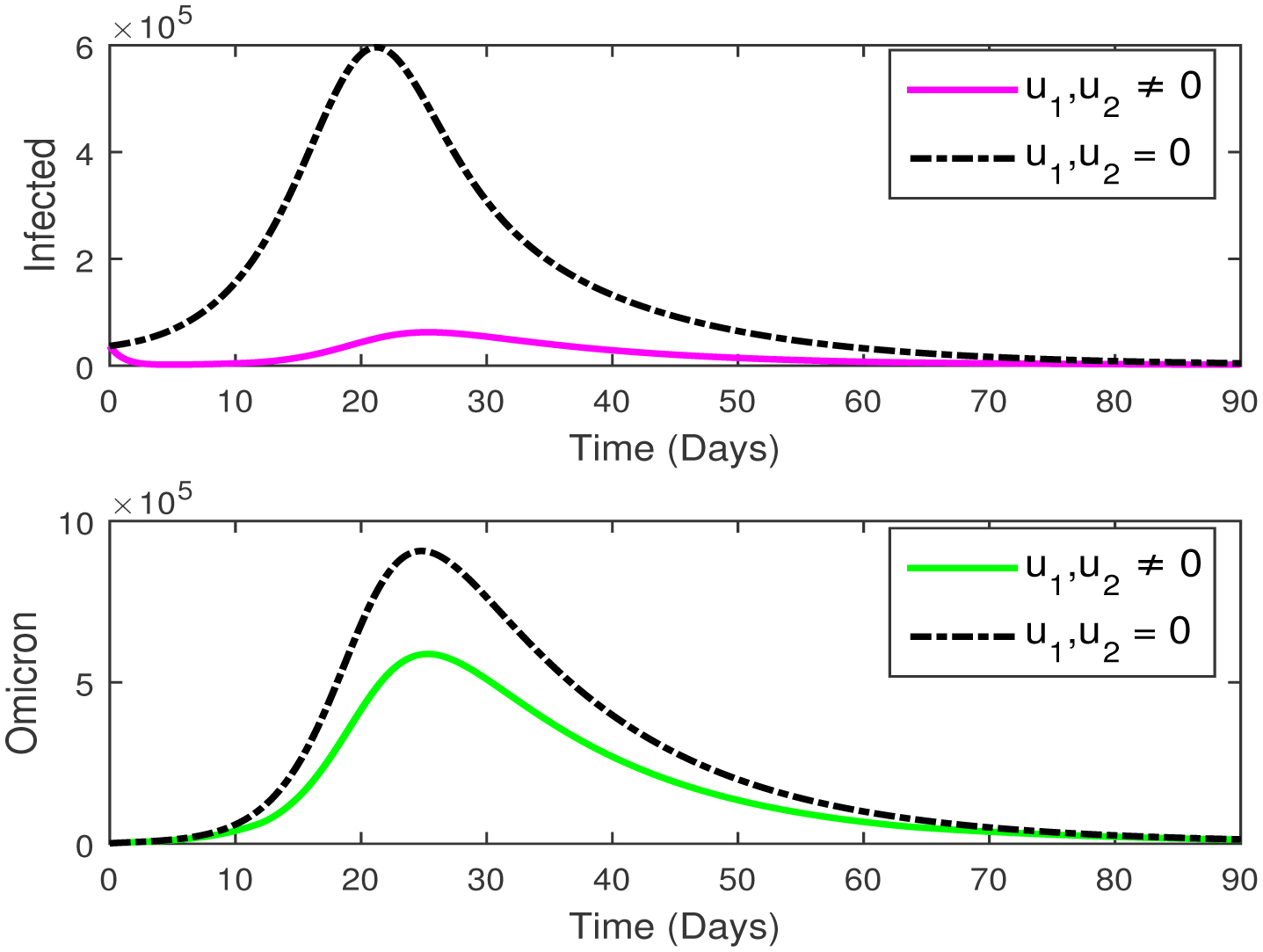
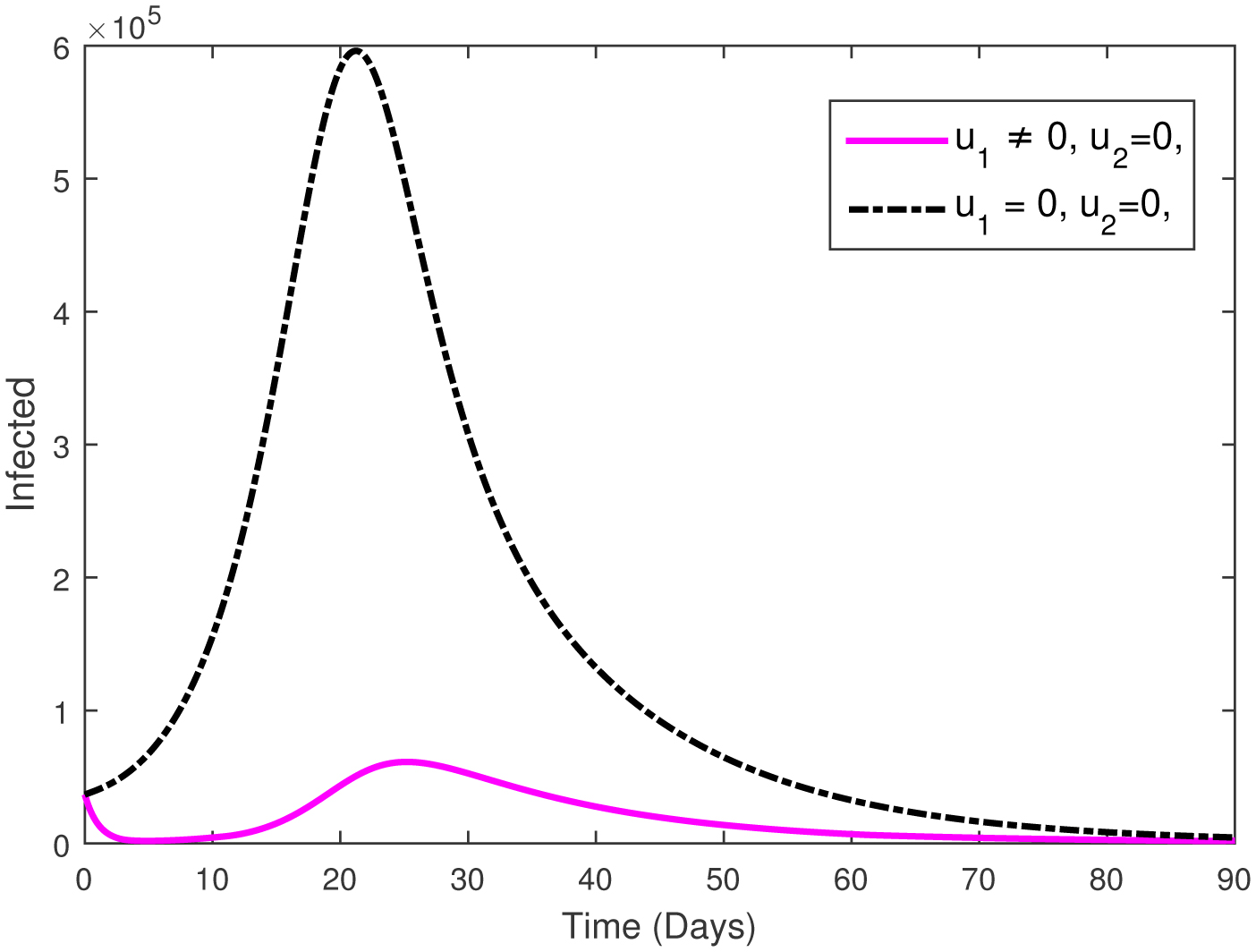
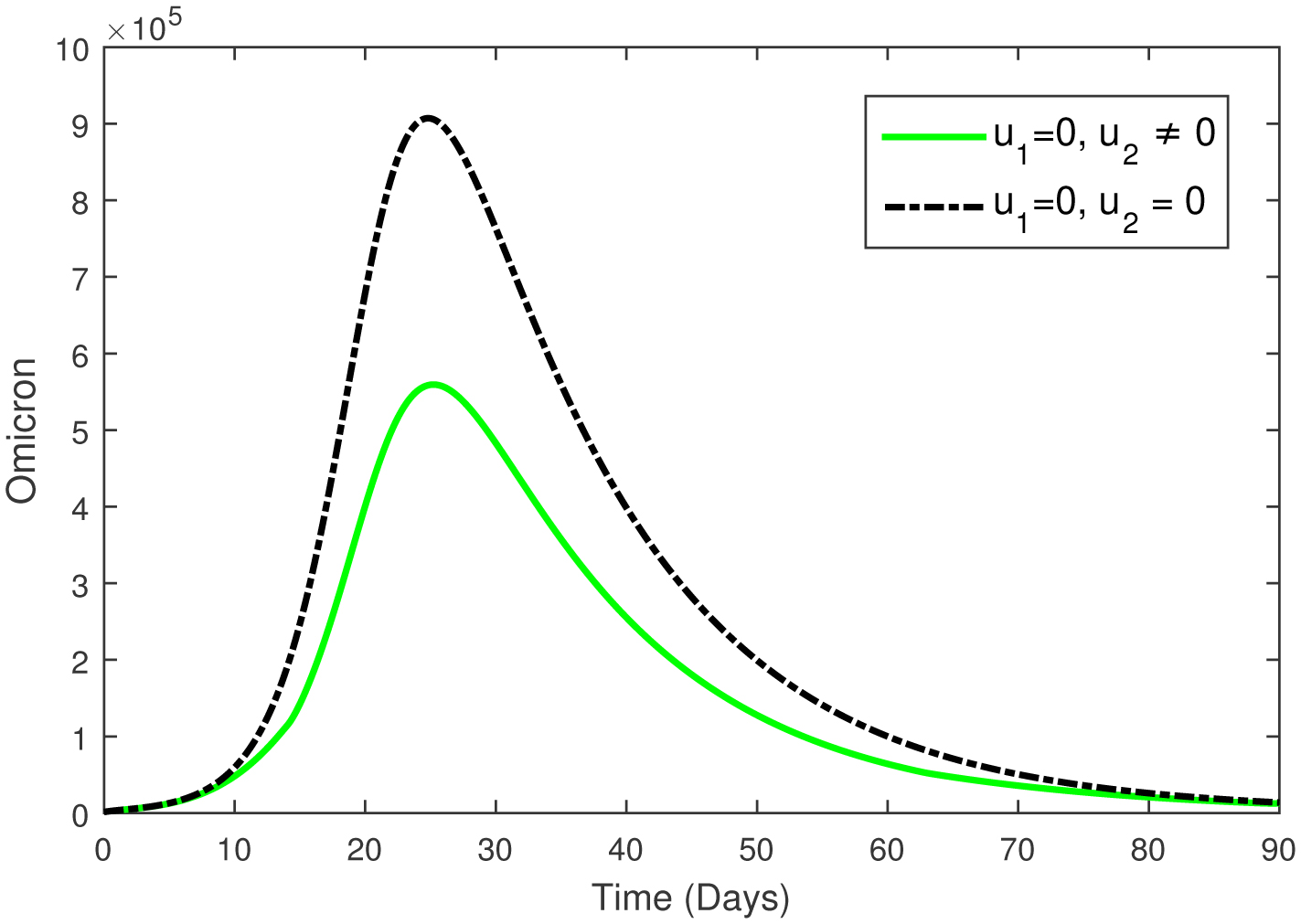
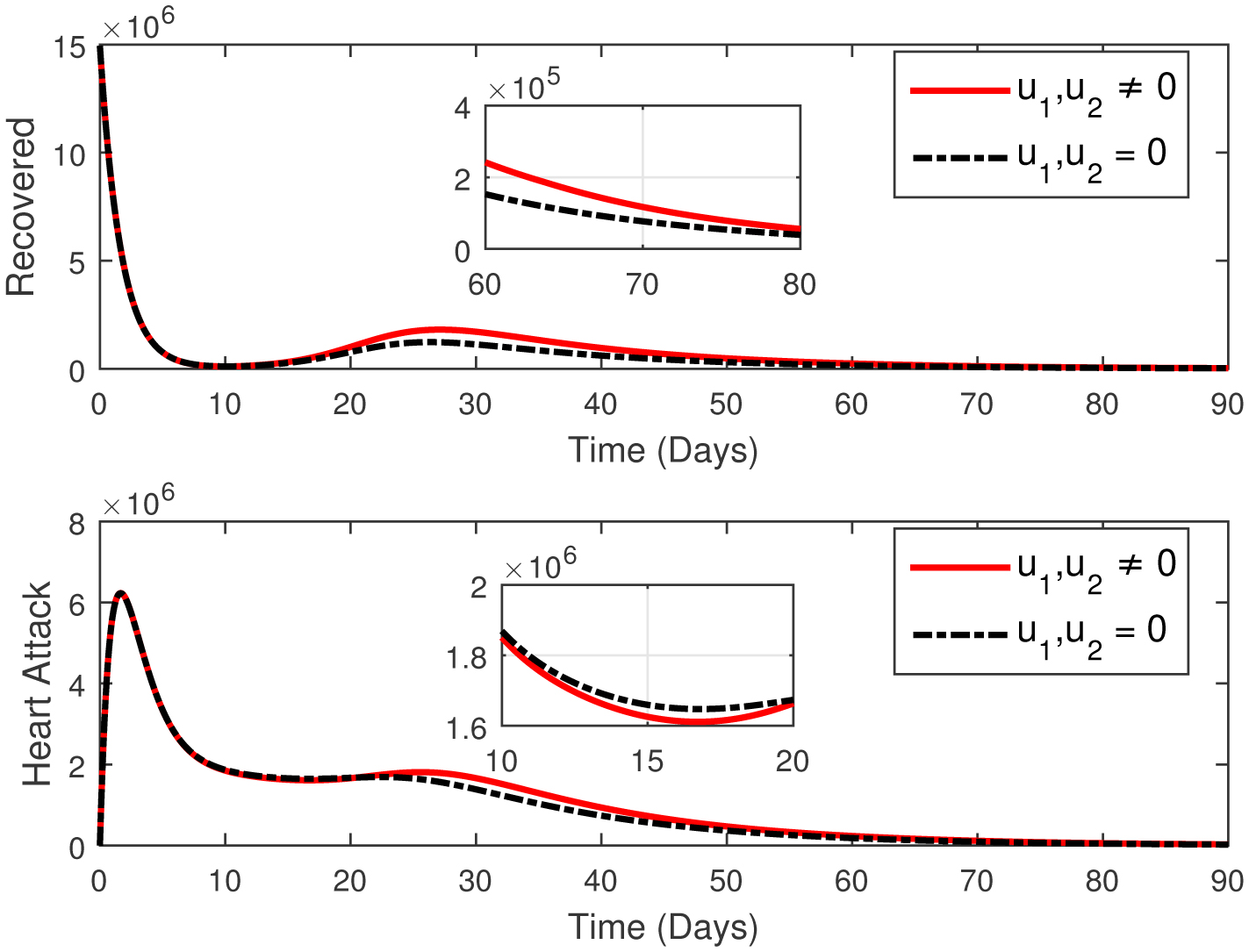
This paper presents an investigation into the relationship between heart attacks and the Omicron variant, employing a novel mathematical model. The model incorporates two adjustable control parameters to manage the number of infected individuals and individuals with the Omicron variant. The study examines the model's positivity and boundedness, evaluates the reproduction number (R0), and conducts a sensitivity analysis of the control parameters based on the reproduction number. The model's parameters are estimated using the widely utilized least squares curve fitting method, employing real COVID-19 cases from Türkiye. Finally, numerical simulations demonstrate the efficacy of the suggested controls in reducing the number of infected individuals and the Omicron population.
Citation: Fırat Evirgen, Fatma Özköse, Mehmet Yavuz, Necati Özdemir. Real data-based optimal control strategies for assessing the impact of the Omicron variant on heart attacks[J]. AIMS Bioengineering, 2023, 10(3): 218-239. doi: 10.3934/bioeng.2023015
This paper presents an investigation into the relationship between heart attacks and the Omicron variant, employing a novel mathematical model. The model incorporates two adjustable control parameters to manage the number of infected individuals and individuals with the Omicron variant. The study examines the model's positivity and boundedness, evaluates the reproduction number (R0), and conducts a sensitivity analysis of the control parameters based on the reproduction number. The model's parameters are estimated using the widely utilized least squares curve fitting method, employing real COVID-19 cases from Türkiye. Finally, numerical simulations demonstrate the efficacy of the suggested controls in reducing the number of infected individuals and the Omicron population.

| [1] | Mishra P, Parveen R, Bajpai R, et al. (2021) Impact of cardiovascular diseases on severity of COVID-19 patients: a systematic review. Ann Acad Med Singap 50: 52-60. https://doi.org/10.47102/annals-acadmedsg.2020367 |
| [2] | Harrison SL, Buckley BJ, Rivera-Caravaca JM, et al. (2021) Cardiovascular risk factors, cardiovascular disease, and COVID-19: an umbrella review of systematic reviews. Eur Heart J-Qual Car 7: 330-339. https://doi.org/10.1093/ehjqcco/qcab029 |
| [3] | Clerkin KJ, Fried JA, Raikhelkar J, et al. (2021) COVID-19 and cardiovascular disease. Circulation 141: 1648-1655. https://doi.org/10.1161/CIRCULATIONAHA.120.046941 |
| [4] | Naik PA, Eskandari Z, Yavuz M, et al. (2022) Complex dynamics of a discrete-time Bazykin-Berezovskaya prey-predator model with a strong Allee effect. J Comput Appl Math 413: 114401. https://doi.org/10.1016/j.cam.2022.114401 |
| [5] | Sene N (2022) Second-grade fluid with Newtonian heating under Caputo fractional derivative: analytical investigations via Laplace transforms. Math Model Num Simul Appl 2: 13-25. https://doi.org/10.53391/mmnsa.2022.01.002 |
| [6] | Sabbar Y (2023) Asymptotic extinction and persistence of a perturbed epidemic model with different intervention measures and standard lévy jumps. Bull Biomath 1: 58-77. https://doi.org/10.59292/bulletinbiomath.2023004 |
| [7] | Hammouch Z, Yavuz M, Özdemir N (2021) Numerical solutions and synchronization of a variable-order fractional chaotic system. Math Model Num Simul Appl 1: 11-23. https://doi.org/10.53391/mmnsa.2021.01.002 |
| [8] | Naik PA, Yavuz M, Qureshi S, et al. (2020) Modeling and analysis of COVID-19 epidemics with treatment in fractional derivatives using real data from Pakistan. Eur Phys J Plus 135: 1-42. https://doi.org/10.1140/epjp/s13360-020-00819-5 |
| [9] | Joshi H, Yavuz M, Townley S, et al. (2023) Stability analysis of a non-singular fractional-order covid-19 model with nonlinear incidence and treatment rate. Phys Scripta 98: 045216. https://doi.org/10.1088/1402-4896/acbe7a |
| [10] | Atede AO, Omame A, Inyama SC (2023) A fractional order vaccination model for COVID-19 incorporating environmental transmission: a case study using Nigerian data. Bull Biomath 1: 78-110. https://doi.org/10.59292/bulletinbiomath.2023005 |
| [11] | Uçar S, Uçar E, Özdemir N, et al. (2019) Mathematical analysis and numerical simulation for a smoking model with Atangana–Baleanu derivative. Chaos Soliton Fract 118: 300-306. https://doi.org/10.1016/j.chaos.2018.12.003 |
| [12] | Naik PA, Owolabi KM, Yavuz M, et al. (2020) Chaotic dynamics of a fractional order HIV-1 model involving AIDS-related cancer cells. Chaos Soliton Fract 140: 110272. https://doi.org/10.1016/j.chaos.2020.110272 |
| [13] | Evirgen F, Ucar E, Ucar S, et al. (2023) Modelling influenza a disease dynamics under Caputo-Fabrizio fractional derivative with distinct contact rates. Math Model Num Simul Appl 3: 58-72. https://doi.org/10.53391/mmnsa.1274004 |
| [14] | Uçar E, Uçar S, Evirgen F, et al. (2021) A fractional SAIDR model in the frame of Atangana–Baleanu derivative. Fractal Fract 5: 32. https://doi.org/10.3390/fractalfract5020032 |
| [15] | Elhia M, Balatif O, Boujallal L, et al. (2021) Optimal control problem for a tuberculosis model with multiple infectious compartments and time delays. IJOCTA 11: 75-91. https://doi.org/10.11121/ijocta.01.2021.00885 |
| [16] | Nwajeri UK, Atede AO, Panle AB, et al. (2023) Malaria and cholera co-dynamic model analysis furnished with fractional-order differential equations. Math Model Num Simul Appl 3: 33-57. https://doi.org/10.53391/mmnsa.1273982 |
| [17] | Agarwal P, Nieto JJ, Torres DFM (2022) Mathematical Analysis of Infectious Diseases. Academic Press. https://doi.org/10.1016/C2020-0-03443-2 |
| [18] | Yıldız TA, Arshad S, Baleanu D (2018) New observations on optimal cancer treatments for a fractional tumor growth model with and without singular kernel. Chaos Soliton Fract 117: 226-239. https://doi.org/10.1016/j.chaos.2018.10.029 |
| [19] | Malinzi J, Ouifki R, Eladdadi A, et al. (2018) Enhancement of chemotherapy using oncolytic virotherapy: Mathematical and optimal control analysis. Math Biosci Eng 15: 1435-1463. https://doi.org/10.3934/mbe.2018066 |
| [20] | Yıldız TA (2019) A comparison of some control strategies for a non-integer order tuberculosis model. IJOCTA 9: 21-30. https://doi.org/10.11121/ijocta.01.2019.00657 |
| [21] | Yıldız TA, Karaoğlu E (2019) Optimal control strategies for tuberculosis dynamics with exogenous reinfections in case of treatment at home and treatment in hospital. Nonlinear Dynam 97: 2643-2659. https://doi.org/10.1007/s11071-019-05153-9 |
| [22] | Baleanu D, Jajarmi A, Sajjadi SS, et al. (2019) A new fractional model and optimal control of a tumor-immune surveillance with non-singular derivative operator. Chaos 29: 083127. https://doi.org/10.1063/1.5096159 |
| [23] | Abidemi A, Aziz NAB (2020) Optimal control strategies for dengue fever spread in Johor, Malaysia. Comput Meth Prog Bio 196: 105585. https://doi.org/10.1016/j.cmpb.2020.105585 |
| [24] | Jajarmi A, Yusuf A, Baleanu D, et al. (2020) A new fractional HRSV model and its optimal control: a non-singular operator approach. Physica A 547: 123860. https://doi.org/10.1016/j.physa.2019.123860 |
| [25] | Naik PA, Zu J, Owolabi KM (2020) Global dynamics of a fractional order model for the transmission of HIV epidemic with optimal control. Chaos Soliton Fract 138: 109826. https://doi.org/10.1016/j.chaos.2020.109826 |
| [26] | Ameen I, Baleanu D, Ali HM (2020) An efficient algorithm for solving the fractional optimal control of SIRV epidemic model with a combination of vaccination and treatment. Chaos Soliton Fract 137: 109892. https://doi.org/10.1016/j.chaos.2020.109892 |
| [27] | Elhia M, Balatif O, Boujallal L, et al. (2021) Optimal control problem for a tuberculosis model with multiple infectious compartments and time delays. IJOCTA 11: 75-91. https://doi.org/10.11121/ijocta.01.2021.00885 |
| [28] | Sweilam NH, Al-Mekhlafi SM, Albalawi AO, et al. (2021) Optimal control of variable-order fractional model for delay cancer treatments. Appl Math Model 89: 1557-1574. https://doi.org/10.1016/j.apm.2020.08.012 |
| [29] | Zhao J, Yang R (2021) A dynamical model of echinococcosis with optimal control and cost-effectiveness. Nonlinear Anal Real World Appl 62: 103388. https://doi.org/10.1016/j.nonrwa.2021.103388 |
| [30] | Mohammadi H, Kumar S, Rezapour S, et al. (2021) A theoretical study of the Caputo–Fabrizio fractional modeling for hearing loss due to Mumps virus with optimal control. Chaos Soliton Fract 144: 110668. https://doi.org/10.1016/j.chaos.2021.110668 |
| [31] | Abbasi Z, Zamani I, Mehra AHA, et al. (2020) Optimal control design of impulsive SQEIAR epidemic models with application to COVID-19. Chaos Soliton Fract 139: 110054. https://doi.org/10.1016/j.chaos.2020.110054 |
| [32] | Moussouni N, Aliane M (2021) Optimal control of COVID-19. IJOCTA 11: 114-122. https://doi.org/10.11121/ijocta.01.2021.00974 |
| [33] | Nabi KN, Kumar P, Erturk VS (2021) Projections and fractional dynamics of COVID-19 with optimal control strategies. Chaos Soliton Fract 145: 110689. https://doi.org/10.1016/j.chaos.2021.110689 |
| [34] | Arruda EF, Das SS, Dias CM, et al. (2021) Modelling and optimal control of multi strain epidemics, with application to COVID-19. Plos One 16: e0257512. https://doi.org/10.1371/journal.pone.0257512 |
| [35] | Tchoumi SY, Diagne ML, Rwezaura H, et al. (2021) Malaria and COVID-19 co-dynamics: A mathematical model and optimal control. Appl Math Model 99: 294-327. https://doi.org/10.1016/j.apm.2021.06.016 |
| [36] | Araz SI (2021) Analysis of a Covid-19 model: optimal control, stability and simulations. Alexandria Eng J 60: 647-658. https://doi.org/10.1016/j.aej.2020.09.058 |
| [37] | Rosa S, Torres DFM (2022) Fractional modelling and optimal control of COVID-19 transmission in Portugal. Axioms 11: 170. https://doi.org/10.3390/axioms11040170 |
| [38] | Fatima B, Yavuz M, ur Rahman M, et al. (2023) Modeling the epidemic trend of middle eastern respiratory syndrome coronavirus with optimal control. Math Biosc Eng 20: 11847-11874. https://doi.org/10.3934/mbe.2023527 |
| [39] | Özköse F, Yavuz M (2022) Investigation of interactions between COVID-19 and diabetes with hereditary traits using real data: a case study in Turkey. Comput Biol Med 141: 105044. https://doi.org/10.1016/j.compbiomed.2021.105044 |
| [40] | Driessche VP, Watmough J (2002) Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math Biosci 180: 29-48. https://doi.org/10.1016/S0025-5564(02)00108-6 |
| [41] | Diekmann O, Heesterbeek JAP, Roberts MG (2010) The construction of next-generation matrices for compartmental epidemic models. J R Soc Interface 7: 873-885. https://doi.org/10.1098/rsif.2009.0386 |
| [42] | Chitnis N, Hyman JM, Cushing JM (2008) Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model. Bull Math Biol 70: 1272. https://doi.org/10.1007/s11538-008-9299-0 |
| [43] | Coddington EA, Levinson N Theory of Ordinary Differential Equations (1955). Tata McGraw-Hill Education |
| [44] | Gaff HD, Schaefer E, Lenhart S (2011) Use of optimal control models to predict treatment time for managing tick-borne disease. J Biol Dynam 5: 517-530. https://doi.org/10.1080/17513758.2010.535910 |
| [45] |
. Pontryagin LS (1987) |
| [46] |
. Lenhart S, Workman JT (2007) |
| [47] | Vitiello A, Ferrara F, Auti AM, et al. (2022) Advances in the Omicron variant development. J Int Med 292: 81-90. https://doi.org/10.1111/joim.13478 |
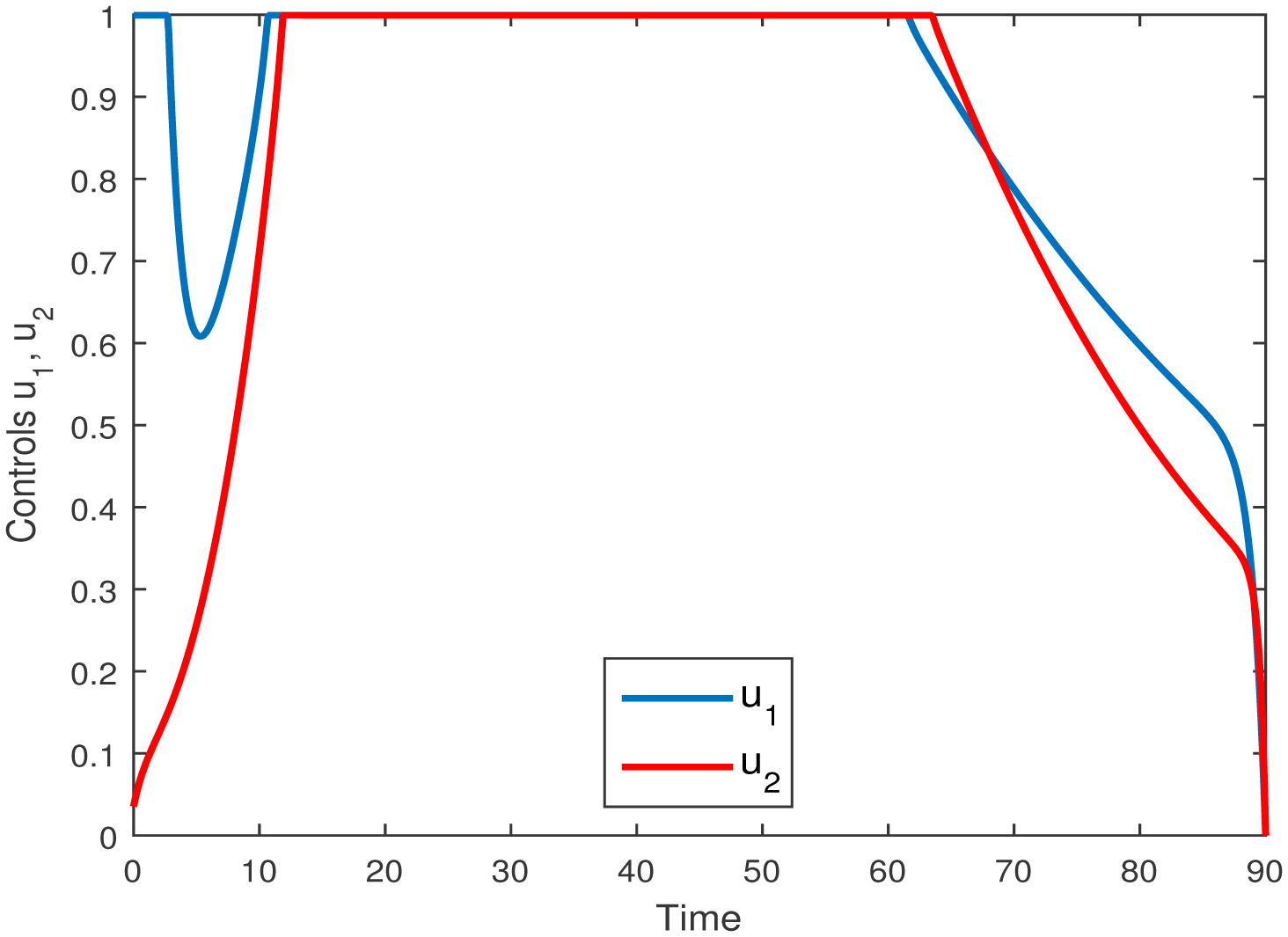
Figures(9) / Tables(3)

Fırat Evirgen, Fatma Özköse, Mehmet Yavuz, Necati Özdemir. Real data-based optimal control strategies for assessing the impact of the Omicron variant on heart attacks[J]. AIMS Bioengineering, 2023, 10(3): 218-239. doi: 10.3934/bioeng.2023015







 DownLoad:
DownLoad: