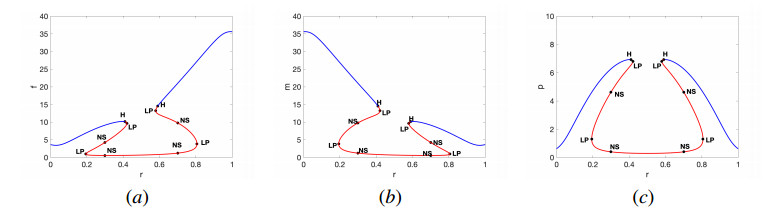
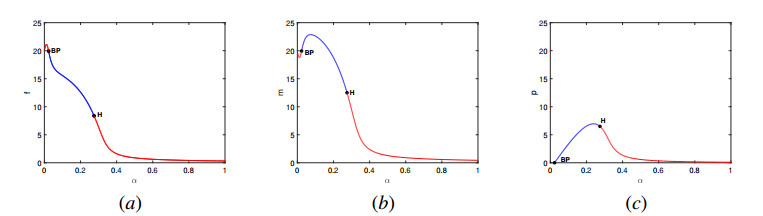
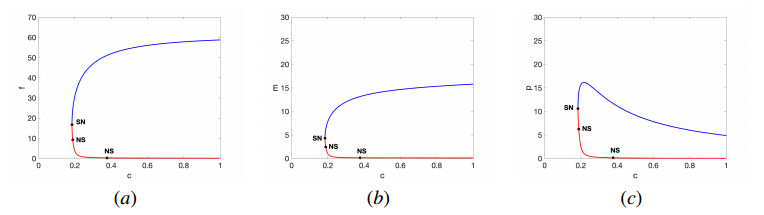
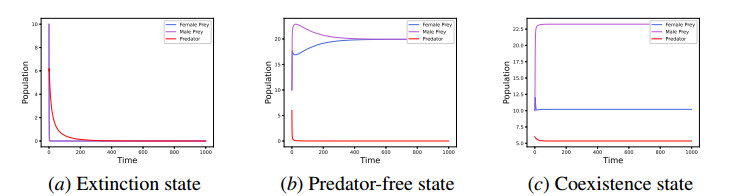
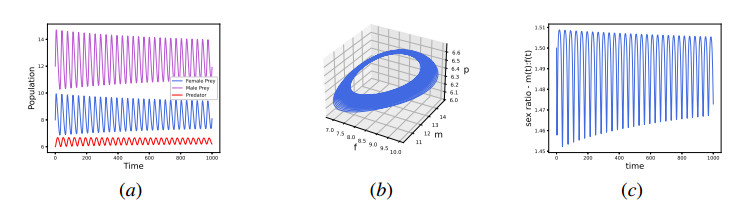
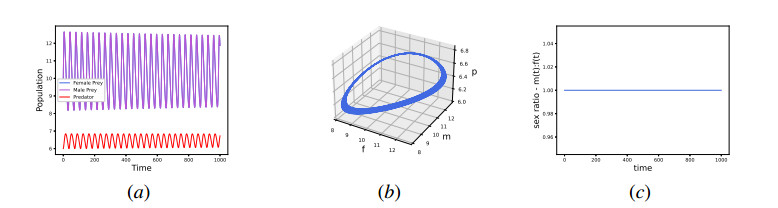
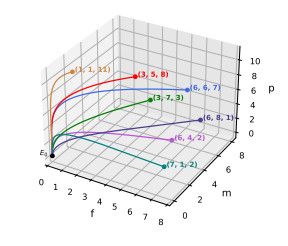
In this work, we propose and investigate a predator-prey model where the prey population is structured by sex and the predators (unstructured) depredate based on sex-bias. We provide conditions for the existence of equilibrium points and perform local stability analysis on them. We derive global stability conditions for the extinction state. We show the possible occurrence of Hopf and saddle-node bifurcations. Multiple Hopf bifurcations are observed as the sex-biased predation rate is varied. This variation also shows the opposite consequences in the densities of the sex-structured prey. Our results show that sex-biased predation can cause both stabilizing and destabilizing effects for certain parameter choices. It can also cause an imbalanced sex-ratio, which has ecological consequences. Furthermore when intraspecific competition among predators is minimized, it can lead to the extinction of prey. We discuss the ecological implications and application of our results to the biocontrol of invasive species susceptible to sex-biased predation.
Citation: Eric M. Takyi, Charles Ohanian, Margaret Cathcart, Nihal Kumar. Sex-biased predation and predator intraspecific competition effects in a prey mating system[J]. AIMS Mathematics, 2024, 9(1): 2435-2453. doi: 10.3934/math.2024120
In this work, we propose and investigate a predator-prey model where the prey population is structured by sex and the predators (unstructured) depredate based on sex-bias. We provide conditions for the existence of equilibrium points and perform local stability analysis on them. We derive global stability conditions for the extinction state. We show the possible occurrence of Hopf and saddle-node bifurcations. Multiple Hopf bifurcations are observed as the sex-biased predation rate is varied. This variation also shows the opposite consequences in the densities of the sex-structured prey. Our results show that sex-biased predation can cause both stabilizing and destabilizing effects for certain parameter choices. It can also cause an imbalanced sex-ratio, which has ecological consequences. Furthermore when intraspecific competition among predators is minimized, it can lead to the extinction of prey. We discuss the ecological implications and application of our results to the biocontrol of invasive species susceptible to sex-biased predation.

| [1] |
E. M. Takyi, K. Cooper, A. Dreher, C. McCrorey, Dynamics of a predator–prey system with wind effect and prey refuge, Journal of Applied Nonlinear Dynamics, 12 (2023), 427–440. https://doi.org/10.5890/JAND.2023.09.001 doi: 10.5890/JAND.2023.09.001

|
| [2] |
E. M. Takyi, K. Antwi-Fordjour, Counter-attack in a stage-structured model with adult predator-induced fear and competition. International Journal of Dynamics and Control, (2023), 1–13. https://doi.org/10.1007/s40435-023-01193-7 doi: 10.1007/s40435-023-01193-7
|
| [3] |
K. Antwi-Fordjour, R. D. Parshad, M. A. Beauregard, Dynamics of a predator–prey model with generalized Holling type functional response and mutual interference, Math. Biosci., 326 (2020), 108407. https://doi.org/10.1016/j.mbs.2020.108407 doi: 10.1016/j.mbs.2020.108407
|
| [4] |
R. D. Parshad, S. Wickramasooriya, K. Antwi-Fordjour, A. Banerjee, Additional food causes predators to explode-unless the predators compete, International Journal of Bifurcation and Chaos, 33 (2013), 2350034. https://doi.org/10.1142/S0218127423500347 doi: 10.1142/S0218127423500347
|
| [5] |
S. A. Jose, R. Ramachandran, J. Cao, J. Alzabut, M. Niezabitowski, V. E. Balas, Stability analysis and comparative study on different eco-epidemiological models: stage structure for prey and predator concerning impulsive control, Optim. Contr. Appl. Met., 43 (2022), 842–866. https://doi.org/10.1002/oca.2856 doi: 10.1002/oca.2856
|
| [6] |
S. R. Hoy, S. J. Petty, A. Millon, D. P. Whitfield, M. Marquiss, M. Davison, et al., Age and sex-selective predation moderate the overall impact of predators, J. Anim. Ecol., 84 (2015), 692–701. https://doi.org/10.1111/1365-2656.12310 doi: 10.1111/1365-2656.12310
|
| [7] |
W. Xu, J. Zhang, S. Du, Q. Dai, W. Zhang, M. Luo, B. Zhao, Sex differences in alarm response and predation risk in the fresh water snail Pomacea canaliculata, J. Mollus. Stud., 80 (2014), 117–122. https://doi.org/10.1093/mollus/eyt054 doi: 10.1093/mollus/eyt054
|
| [8] |
N. Skals, D. Plepys, C. Löfstedt, Foraging and mate-finding in the silver Y moth, Autographa gamma (Lepidoptera: Noctuidae) under the risk of predation, Oikos, 102 (2003), 351–357. https://doi.org/10.1034/j.1600-0706.2003.12627.x doi: 10.1034/j.1600-0706.2003.12627.x
|
| [9] | R. Naylor, Invasions in agriculture: assessing the cost of the golden apple snail in Asia, Ambio, (1996), 443–448. |
| [10] | A. L. Estebenet, P. R. Martín, Minireview: Pomacea canaliculata (Gastropoda: Ampullariidae): life-history traits and their plasticity, Biocell, 26 (2002), 83. |
| [11] |
N. O. L. Carlsson, C. Brönmark, L.-A. Hansson, Invading herbivory: the golden apple snail alters ecosystem functioning in Asian wetlands, Ecology, 85 (2004), 1575–1580. https://doi.org/10.1890/03-3146 doi: 10.1890/03-3146
|
| [12] | S. Lowe, M. Browne, S. Boudjelas, M. De Poorter, 100 of the world's worst invasive alien species: a selection from the global invasive species database, volume 12, Invasive Species Specialist Group Auckland, 2000. |
| [13] |
A. E. McKellar, M. M. Turcotte, A. P. Hendry, Environmental factors influencing adult sex ratio in Trinidadian guppies, Oecologia, 159 (2009), 735–745. https://doi.org/10.1007/s00442-008-1257-x doi: 10.1007/s00442-008-1257-x
|
| [14] |
J. L. DeGabriel, B. D. Moore, W. J. Foley, C. N. Johnson, Male-biased predation and its effect on paternity skew and life history in a population of common brushtail possums (Trichosurus vulpecula), PLoS One, 9 (2014), e111746. https://doi.org/10.1371/journal.pone.0111746 doi: 10.1371/journal.pone.0111746
|
| [15] |
J. Gao, F. Santi, L. Zhou, X. Wang, R. Riesch, M. Plath, Geographical and temporal variation of multiple paternity in invasive mosquitofish (Gambusia holbrooki, Gambusia affinis), Mol. Ecol., 28 (2019), 5315–5329. https://doi.org/10.1111/mec.15294 doi: 10.1111/mec.15294
|
| [16] |
R. H. Britton, M. E. Moser, Size specific predation by herons and its effect on the sex-ratio of natural populations of the mosquito fish Gambusia affinis baird and girard, Oecologia, 53 (1982), 146–151. https://doi.org/10.1007/BF00545657 doi: 10.1007/BF00545657
|
| [17] |
J. G. J. Godin, Predation risk and alternative mating tactics in male Trinidadian guppies (Poecilia reticulata), Oecologia, 103 (1995), 224–229. https://doi.org/10.1007/BF00329084 doi: 10.1007/BF00329084
|
| [18] |
B. B. M. Wong, P. A. Svensson, Strategic male signalling effort in a desert-dwelling fish, Behav. Ecol. Sociobiol., 63 (2009), 543–549. https://doi.org/10.1007/s00265-008-0689-y doi: 10.1007/s00265-008-0689-y
|
| [19] |
T. K. Lehtonen, P. A. Svensson, B. Wong, Both male and female identity influence variation in male signalling effort, BMC Evol. Biol., 11 (2011), 1–8. https://doi.org/10.1186/1471-2148-11-233 doi: 10.1186/1471-2148-11-233
|
| [20] |
R. Heinsohn, J. Au, H. Kokko, M. H. Webb, R. M. Deans, R. Crates, et al., Can an introduced predator select for adaptive sex allocation? Proceedings of the Royal Society B, 288 (1949), 20210093. https://doi.org/10.1098/rspb.2021.0093 doi: 10.1098/rspb.2021.0093
|
| [21] |
T. Székely, F. J. Weissing, J. Komdeur, Adult sex ratio variation: implications for breeding system evolution, J. Evolution. Biol., 27 (2014), 1500–1512. https://doi.org/10.1111/jeb.12415 doi: 10.1111/jeb.12415
|
| [22] |
D. Bierbach, M. Schulte, N. Herrmann, M. Tobler, S. Stadler, C. T. Jung, et al., Predator-induced changes of female mating preferences: innate and experiential effects, BMC Evol. Biol., 11 (2011), 1–11. https://doi.org/10.1186/1471-2148-11-190 doi: 10.1186/1471-2148-11-190
|
| [23] |
T. Lodé, M.-J. Holveck, D. Lesbarreres, A. Pagano, Sex–biased predation by polecats influences the mating system of frogs, Proceedings of the Royal Society of London. Series B: Biological Sciences, 271(suppl_6) (2004), S399–S401. https://doi.org/10.1098/rsbl.2004.0195 doi: 10.1098/rsbl.2004.0195
|
| [24] |
D. S. Boukal, L. Berec, V. Křivan, Does sex-selective predation stabilize or destabilize predator-prey dynamics? PLoS One, 3 (2008), e2687. https://doi.org/10.1371/journal.pone.0002687 doi: 10.1371/journal.pone.0002687
|
| [25] |
D. I. Bolnick, Can intraspecific competition drive disruptive selection? An experimental test in natural populations of sticklebacks, Evolution, 58 (2004), 608–618. https://doi.org/10.1111/j.0014-3820.2004.tb01683.x doi: 10.1111/j.0014-3820.2004.tb01683.x
|
| [26] |
P. N. Mwangi, M. Schmitz, C. Scherber, C. Roscher, J. Schumacher, M. Scherer-Lorenzen, et al., Niche pre-emption increases with species richness in experimental plant communities, J. Ecol., 95 (2007), 65–78. https://doi.org/10.1111/j.1365-2745.2006.01189.x doi: 10.1111/j.1365-2745.2006.01189.x
|
| [27] |
J. HilleRisLambers, P. B. Adler, W. S. Harpole, J. M. Levine, M. M. Mayfield, Rethinking community assembly through the lens of coexistence theory, Annual review of Ecology, Evolution, and Systematics, 43 (2012), 227–248. https://doi.org/10.1146/annurev-ecolsys-110411-160411 doi: 10.1146/annurev-ecolsys-110411-160411
|
| [28] |
E. W. Seabloom, W. S. Harpole, O. J. Reichman, D. Tilman, Invasion, competitive dominance, and resource use by exotic and native California grassland species, Proceedings of the National Academy of Sciences, 100 (2003), 13384–13389. https://doi.org/10.1073/pnas.1835728100 doi: 10.1073/pnas.1835728100
|
| [29] |
J. L. Teem, J. B. Gutierrez, R. D. Parshad, A comparison of the Trojan Y chromosome and daughterless carp eradication strategies, Biol. Invasions, 16 (2014), 1217–1230. https://doi.org/10.1007/s10530-013-0475-2 doi: 10.1007/s10530-013-0475-2
|
| [30] |
M. Pierre, Global existence in reaction-diffusion systems with control of mass: a survey, Milan J. Math., 78 (2010), 417–455. https://doi.org/10.1007/s00032-010-0133-4 doi: 10.1007/s00032-010-0133-4
|
| [31] |
M. Pierre, D. Schmitt, Blowup in reaction-diffusion systems with dissipation of mass, SIAM Rev., 42 (2000), 93–106. https://doi.org/10.1137/S0036144599359735 doi: 10.1137/S0036144599359735
|
| [32] | Jingjing Lyu. Mathematical methods in invasive species control, Clarkson University, 2018. |
| [33] |
J. Lyu, P. J. Schofield, K. M. Reaver, M. Beauregard, R. D. Parshad, A comparison of the Trojan Y chromosome strategy to harvesting models for eradication of nonnative species, Nat. Resour. Model., 33 (2020), e12252. https://doi.org/10.1111/nrm.12252 doi: 10.1111/nrm.12252
|
| [34] | L. Perko, Differential equations and dynamical systems, volume 7, Springer Science & Business Media, Springer-Verlag, New York, 2013. |
| [35] |
A. Dhooge, W. Govaerts, Y. A. Kuznetsov, H. Gaétan Ellart Meijer, B. Sautois, New features of the software matcont for bifurcation analysis of dynamical systems, Math. Comput. Model. Dyn., 14 (2008), 147–175. https://doi.org/10.1080/13873950701742754 doi: 10.1080/13873950701742754
|
| [36] | S. H. Alonzo, B. C. Sheldon, Population density, social behaviour and sex allocation, Social behaviour: genes ecology and evolution. Cambridge University Press, Cambridge, (2010), 474–488. https://doi.org/10.1017/CBO9780511781360.037 |
| [37] |
C. Bessa-Gomes, S. Legendre, J. Clobert, Allee effects, mating systems and the extinction risk in populations with two sexes, Ecol. Lett., 7 (2004), 802–812. https://doi.org/10.1111/j.1461-0248.2004.00632.x doi: 10.1111/j.1461-0248.2004.00632.x
|
| [38] |
P. F. Donald, Adult sex ratios in wild bird populations, Ibis, 149 (2007), 671–692. https://doi.org/10.1111/j.1474-919X.2007.00724.x doi: 10.1111/j.1474-919X.2007.00724.x
|
| [39] |
S. Veran, S. R. Beissinger, Demographic origins of skewed operational and adult sex ratios: perturbation analyses of two-sex models, Ecol. Lett., 12 (2009), 129–143. https://doi.org/10.1111/j.1461-0248.2008.01268.x doi: 10.1111/j.1461-0248.2008.01268.x
|
| [40] |
C. Wedekind, G. Evanno, T. Szekely, M. Pompini, O. Darbellay, J. Guthruf, Persistent unequal sex ratio in a population of grayling (salmonidae) and possible role of temperature increase, Conserv. Biol., 27 (2013), 229–234. https://doi.org/10.1111/j.1523-1739.2012.01909.x doi: 10.1111/j.1523-1739.2012.01909.x
|
| [41] |
C. J. A. Bradshaw, A. J. Hoskins, P. J. Haubrock, R. N. Cuthbert, C. Diagne, B. Leroy, et al., Detailed assessment of the reported economic costs of invasive species in Australia, NeoBiota, 67 (2021), 511–550. https://doi.org/10.3897/neobiota.67.58834 doi: 10.3897/neobiota.67.58834
|
| [42] |
J. E. Fantle-Lepczyk, P. J. Haubrock, A. M. Kramer, R. N. Cuthbert, A. J. Turbelin, R. Crystal-Ornelas, et al., Economic costs of biological invasions in the United States, Sci. Total Environ., 806 (2022), 151318. https://doi.org/10.1016/j.scitotenv.2021.151318 doi: 10.1016/j.scitotenv.2021.151318
|
| [43] |
C. Marangi, F. Casella, F. Diele, D. Lacitignola, A. Martiradonna, A. Provenzale, et al., Mathematical tools for controlling invasive species in protected areas, Mathematical Approach to Climate Change and its Impacts: MAC2I, (2020), 211–237. https://doi.org/10.1007/978-3-030-38669-6_8 doi: 10.1007/978-3-030-38669-6_8
|
| [44] |
D. Joseph, R. Ramachandran, J. Alzabut, S. A. Jose, H. Khan, A fractional-order density-dependent mathematical model to find the better strain of Wolbachia, Symmetry, 15 (2023), 845. https://doi.org/10.3390/sym15040845 doi: 10.3390/sym15040845
|
| [45] |
R. Shine, Ecological causes for the evolution of sexual dimorphism: a review of the evidence, The Quarterly review of Biology, 64 (1989), 419–461. https://doi.org/10.1086/416458 doi: 10.1086/416458
|
Figures(7) / Tables(1)

Eric M. Takyi, Charles Ohanian, Margaret Cathcart, Nihal Kumar. Sex-biased predation and predator intraspecific competition effects in a prey mating system[J]. AIMS Mathematics, 2024, 9(1): 2435-2453. doi: 10.3934/math.2024120







 DownLoad:
DownLoad: