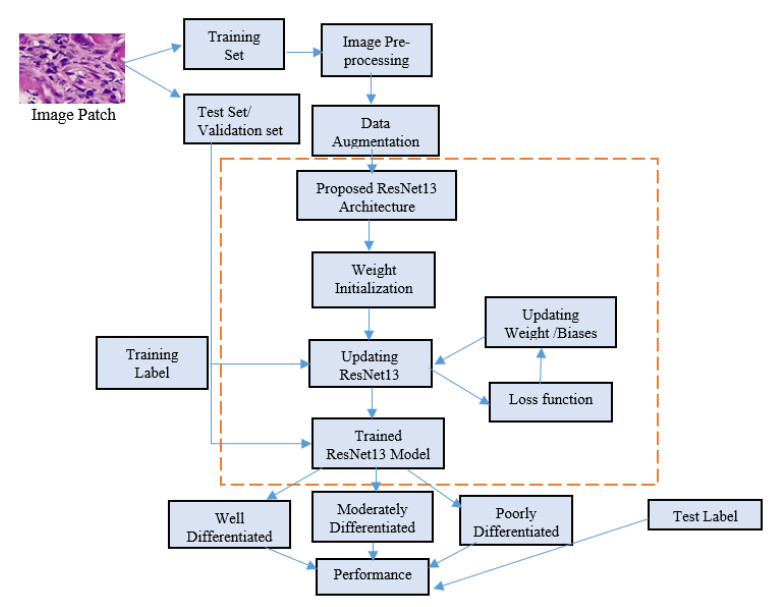
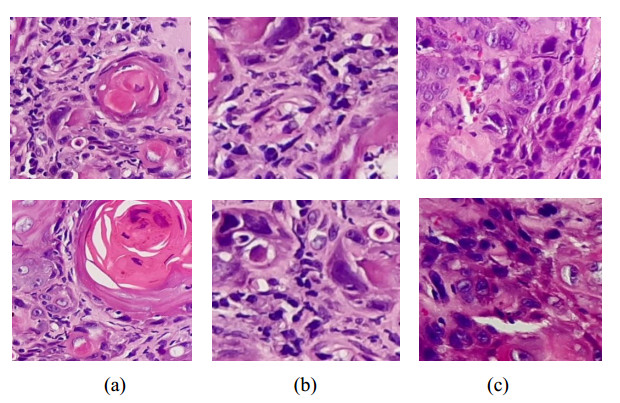
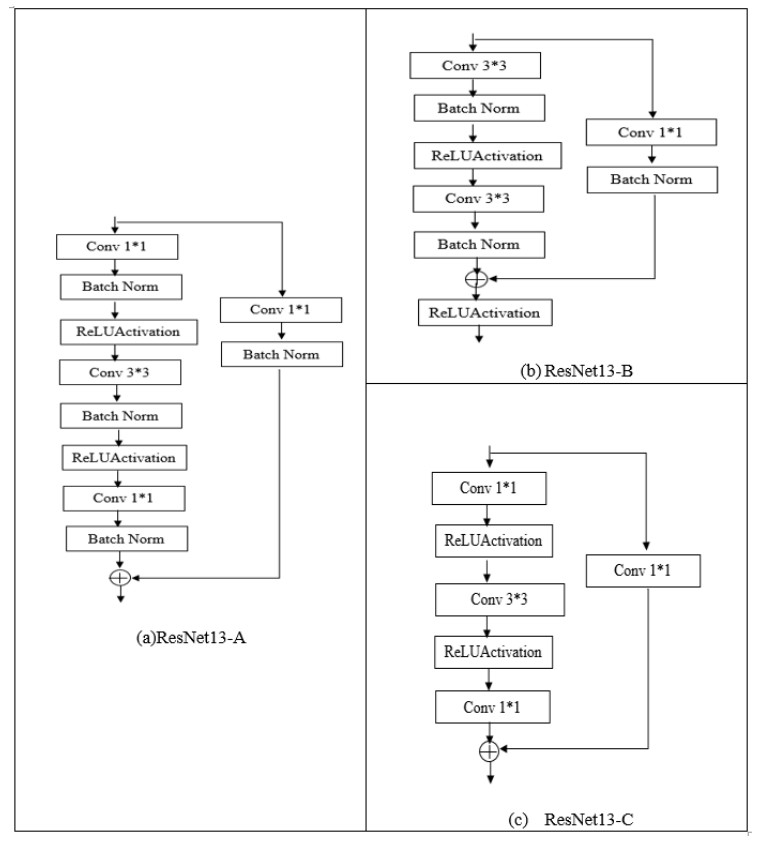
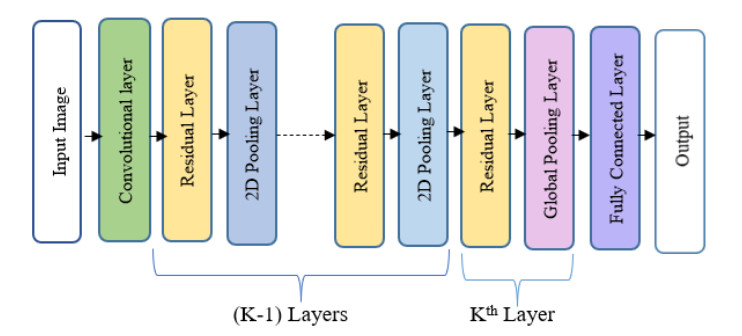
Oral cancer is a prevalent disease happening in the head and neck region. Due to the high occurrence rate and serious consequences of oral cancer, an accurate diagnosis of malignant oral tumors is a major priority. Thus, early diagnosis is very effective to give the patient a prompt response to treatment. The most efficient way for diagnosing oral cancer is from histopathological imaging, which provides a detailed view of inside cells. Accurate and automatic classification of oral histopathological images remains a difficult task due to the complex nature of cell images, staining methods, and imaging conditions. The use of deep learning in imaging techniques and computational diagnostics can assist doctors and physicians in automatically analysing Oral Squamous Cell Carcinoma biopsy images in a timely and efficient manner. Thus, it reduces the operational workload of the pathologist and enhance patient management. Training deeper neural networks takes considerable time and requires a lot of computing resources, due to the complexity of the network and the gradient diffusion problem. With this motivation and inspired by ResNet's significant successes to handle the gradient diffusion problem, in this study we suggest the novel improved ResNet-based model for the automated multistage classification of oral histopathology images. Three prospective candidate model blocks are presented, analyzed, and the best candidate model is chosen as the optimal one which can efficiently classify the oral lesions into well-differentiated, moderately-differentiated and poorly-differentiated in significantly reduced time, with 97.59% accuracy.
Citation: Santisudha Panigrahi, Ruchi Bhuyan, Kundan Kumar, Janmenjoy Nayak, Tripti Swarnkar. Multistage classification of oral histopathological images using improved residual network[J]. Mathematical Biosciences and Engineering, 2022, 19(2): 1909-1925. doi: 10.3934/mbe.2022090
Oral cancer is a prevalent disease happening in the head and neck region. Due to the high occurrence rate and serious consequences of oral cancer, an accurate diagnosis of malignant oral tumors is a major priority. Thus, early diagnosis is very effective to give the patient a prompt response to treatment. The most efficient way for diagnosing oral cancer is from histopathological imaging, which provides a detailed view of inside cells. Accurate and automatic classification of oral histopathological images remains a difficult task due to the complex nature of cell images, staining methods, and imaging conditions. The use of deep learning in imaging techniques and computational diagnostics can assist doctors and physicians in automatically analysing Oral Squamous Cell Carcinoma biopsy images in a timely and efficient manner. Thus, it reduces the operational workload of the pathologist and enhance patient management. Training deeper neural networks takes considerable time and requires a lot of computing resources, due to the complexity of the network and the gradient diffusion problem. With this motivation and inspired by ResNet's significant successes to handle the gradient diffusion problem, in this study we suggest the novel improved ResNet-based model for the automated multistage classification of oral histopathology images. Three prospective candidate model blocks are presented, analyzed, and the best candidate model is chosen as the optimal one which can efficiently classify the oral lesions into well-differentiated, moderately-differentiated and poorly-differentiated in significantly reduced time, with 97.59% accuracy.

| [1] | National Institutes of Health, NIH Fact Sheets-Oral Cancer, (2018), 1–2. Available from: http://www.nidcr.nih.gov. |
| [2] |
B. Timuçin, H. H. Yilmaz, Ü. Aydin, M. A. Aydin, M. C. Aksoy, Derya Yildirim, Early diagnosis of oral cancer, J. Int. Med. Res., 38 (2010), 737–749. doi: 10.1177/147323001003800302. doi: 10.1177/147323001003800302

|
| [3] |
M. M. R. Krishnan, V. Venkatraghavan, U. R. Acharya, M. Pal, R. R. Paul, L. C. Min, et al., Automated oral cancer identification using histopathological images: a hybrid feature extraction paradigm, Micron, 43 (2012), 352–364. doi: 10.1016/j.micron.2011.09.016. doi: 10.1016/j.micron.2011.09.016
|
| [4] |
D. D. Kumar, S. Bose, A. K. Maiti, B. Mitra, G. Mukherjee, P. K. Dutta, Automatic identification of clinically relevant regions from oral tissue histological images for oral squamous cell carcinoma diagnosis, Tissue Cell, 53 (2018), 111–119. doi: 10.1016/j.tice.2018.06.004. doi: 10.1016/j.tice.2018.06.004
|
| [5] |
M. N. Gurcan, L. E. Boucheron, A. Can, A. Madabhushi, N. M. Rajpoot, B. Yener, Histopathological image analysis: A review, IEEE Rev. Biomed. Eng., 2 (2009), 147–171. doi: 10.1109/RBME.2009.2034865. doi: 10.1109/RBME.2009.2034865
|
| [6] |
D. Komura, I. Shumpei, Machine learning methods for histopathological image analysis, Comput. Struct. Biotechnol. J., 16 (2018), 34–42. doi: 10.1016/j.csbj.2018.01.001. doi: 10.1016/j.csbj.2018.01.001
|
| [7] | D. Wang, A. Khosla, R. Gargeya, H. Irshad, A. H. Beck, Deep learning for identifying metastatic breast cancer, (2016), arXiv preprint arXiv: 1606.05718. |
| [8] | D. Wang, D. J. Foran, J. Ren, H. Zhong, I. Y. Kim, X. Qi, Exploring automatic prostate histopathology image gleason grading via local structure modelling, in 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), (2015), 2649–2652. doi: 10.1109/EMBC.2015.7318936. |
| [9] | F. Mohd, N. Noor, Z. A. Bakar, Z. A. Rajion, Analysis of oral cancer prediction using features selection with machine learning, in 2015 The 7th International Conference on Information Technology (ICIT), (2015), 283–288. doi: 10.15849/icit.2015.0058. |
| [10] | M. M. R. Krishnan, C. Chakraborty, A. K. Ray, Wavelet based texture classification of oral histopathological sections, Int. J. Microsc. Sci. Technol. Appl. Educ., 2 (2010), 897–906. |
| [11] | R. Patra, C. Chakraborty, J. Chatterjee, Textural analysis of spinous layer for grading oral submucous fibrosis, Int J. Comput Appl., 47 (2012), 975–8887. |
| [12] |
P. H. Montero, S. G. Patel, Cancer of the oral cavity, Surg. Oncol. Clin., 24 (2015), 491–508. doi: 10.1016/j.soc.2015.03.006. doi: 10.1016/j.soc.2015.03.006
|
| [13] |
M. Akhter, S. Hossain, Q. B. Rahman, M.R. Molla, A study on histological grading of oral squamous cell carcinoma and its co-relationship with regional metastasis, J. Oral Maxillofac. Pathol., 15 (2011), 168. doi: 10.4103/0973-029X.84485. doi: 10.4103/0973-029X.84485
|
| [14] |
M. Y. Chen, H. S. Chiang, E. Lughofer, E. Egrioglu, Deep learning: Emerging trends, applications and research challenges. Soft Comput., 24 (2020), 7835–7838. doi: 10.1007/s00500-020-04939-z. doi: 10.1007/s00500-020-04939-z
|
| [15] | A. Krizhevsky, I. Sutskever, G. E. Hinton, Imagenet classification with deep convolutional neural networks, Adv. Neural Inf. Process. Syst., 25 (2012), 1097–1105. |
| [16] |
S. P. Mohanty, D. P. Hughes, M. Salathe, Using deep learning for image-based plant disease detection, Front. Plant Sci., 7 (2016), 1419. doi: 10.3389/fpls.2016.01419. doi: 10.3389/fpls.2016.01419
|
| [17] | S. H. Kassani, P. H. Kassani, M. J. Wesolowski, K. A. Schneider, R. Deters, Classification of histopathological biopsy images using ensemble of deep learning networks, (2019), arXiv preprint arXiv: 1909.11870. |
| [18] | K. Simonyan, A. Zisserman, Very deep convolutional networks for large-scale image recognition, (2014), arXiv preprint arXiv: 1409.1556. |
| [19] | C. Szegedy, W. Liu, Y. Jia, P. Sermanet, S. Reed, D. Anguelov, et al., Going deeper with convolutions, in Proceedings of the IEEE conference on computer vision and pattern recognition, (2015), 1–9. doi: 10.1109/CVPR.2015.7298594. |
| [20] |
D. W. Kim, S. Lee, S. Kwon, W. Nam, I. H. Cha, H. J. Kim, Deep learning-based survival prediction of oral cancer patients, Sci. Rep., 9 (2019), 1–10. doi: 10.1038/s41598-019-43372-7. doi: 10.1038/s41598-019-43372-7
|
| [21] | D. Gaur, J. Folz, A. Dengel, Training Deep Neural Networks Without Batch Normalization, (2020), arXiv preprint arXiv: 2008.07970. |
| [22] |
M. Krishnan, U. Acharya, C. Chakraborty, A. Ray, Automated diagnosis of oral cancer using higher order spectra features and local binary pattern: A comparative study, Technol. Cancer Res. Treat., 10 (2011), 443–455. doi: 10.7785/tcrt.2012.500221. doi: 10.7785/tcrt.2012.500221
|
| [23] | S. K. Prabhakar, H. Rajaguru, Performance analysis of linear layer neural networks for oral cancer classification, in 2017 6th ICT International Student Project Conference (ICT-ISPC), IEEE, (2017), 1–4. doi: 10.1109/ICT-ISPC.2017.8075357. |
| [24] |
T. Rahman, L. Mahanta, C. Chakraborty, A. Das, J. Sarma, Textural pattern classification for oral squamous cell carcinoma, J. Microsc., 269 (2018), 85–93. doi: 10.1111/jmi.12611. doi: 10.1111/jmi.12611
|
| [25] | T. Y. Rahman, L. B. Mahanta, A. K. Das, J. D. Sarma, Automated oral squamous cell carcinoma identification using shape, texture and color features of whole image strips, Tissue Cell, 63 (2020), 101322. |
| [26] |
P. G. Betta, A. Andrion, A. Donna, F. Mollo, M. Scelsi, G. Zai, C. Magnani, Malignant mesothelioma of the pleura. The reproducibility of the immunohistological diagnosis, Pathol. Res. Pract., 193 (1997), 759–765. doi: 10.1016/S03440338(97)80054-4. doi: 10.1016/S03440338(97)80054-4
|
| [27] |
L. Shi, W. Liu, H. Zhang, Y. Xie, D. Wang, A survey of GPU-based medical image computing techniques, Quant. Imaging Med. Surg., 2 (2012), 188. doi: 10.3978/j.issn.2223-4292.2012.08.02. doi: 10.3978/j.issn.2223-4292.2012.08.02
|
| [28] |
C. L. Srinidhi, O. Ciga, A. L. Martel, Deep neural network models for computational histopathology: A survey, Med. Image Anal., 67 (2021), 101813. doi: 10.1016/j.media.2020.101813. doi: 10.1016/j.media.2020.101813
|
| [29] | S. Panigrahi, T. Swarnkar, Automated Classification of Oral Cancer Histopathology images using Convolutional Neural Network, in 2019 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), (2019), 1232–1234. doi: 10.1109/BIBM47256.2019.8982979. |
| [30] | S. Panigrahi, J. Das, T. Swarnkar, Capsule network based analysis of histopathological images of oral squamous cell carcinoma, J. King Saud Univ.-Comput. Inf. Sci., (2020), In press. doi: 10.1016/j.jksuci.2020.11.003. |
| [31] |
O. Driemel, M. Kunkel, M. Hullmann, F. V. Eggeling, U. Müller‐Richter, H. Kosmehl, et al., Diagnosis of oral squamous cell carcinoma and its precursor lesions, JDDG: J. Dtsch. Dermatol. Ges., 5 (2007), 1095–1100. doi: 10.1111/j.1610-0387.2007.06397.x. doi: 10.1111/j.1610-0387.2007.06397.x
|
| [32] |
D. Navarun, E. Hussain, L. B. Mahanta, Automated classification of cells into multiple classes in epithelial tissue of oral squamous cell carcinoma using transfer learning and convolutional neural network, Neural Netw., 128 (2020), 47–60. doi: 10.1016/j.neunet.2020.05.003. doi: 10.1016/j.neunet.2020.05.003
|
| [33] |
L. H. Shehab, O. M. Fahmy, S. M. Gasser, M.S. El-Mhallawy, et al., An efficient brain tumor image segmentation based on deep residual networks (ResNets), J. King Saud Univ. Eng. Sci., 33 (2021), 404–412. doi: 10.1016/j.jksues.2020.06.001. doi: 10.1016/j.jksues.2020.06.001
|
| [34] |
H. Zhu, M. Ma, W. Ma, L. Jiao, S. Hong, J. Shen, et al., A spatial-channel progressive fusion ResNet for remote sensing classification, Inf. Fusion, 70 (2021), 72–87. doi: 10.1016/j.inffus.2020.12.008. doi: 10.1016/j.inffus.2020.12.008
|
| [35] | P. A. Pattanaik, M. Mohit, Z. K. Mohammad, S. N. Panda. Malaria detection using deep residual networks with mobile microscopy, J. King Saud Univ.-Comput. Inf. Sci., (2020), In press. doi: 10.1016/j.jksuci.2020.07.003. |
| [36] |
L. Mohamed, M. Gunasekaran, H. N. T. Mohamed, M. N. K. Eldeen, Fighting against COVID-19: A novel deep learning model based on YOLO-v2 with ResNet-50 for medical face mask detection, Sustain. Cities Soc., 65 (2021), 102600. doi: 10.1016/j.scs.2020.102600. doi: 10.1016/j.scs.2020.102600
|
| [37] |
J. Duan, T. Shi, H. Zhou, J. Xuan, S. Wang, A novel ResNet-based model structure and its applications in machine health monitoring, J. Vib. Control, 27 (2021), 1036–1050. doi: 10.1177/1077546320936506. doi: 10.1177/1077546320936506
|
| [38] | X. Zhang, X. Zhou, M. Lin, J. Sun, Shufflenet: An extremely efficient convolutional neural network for mobile devices, in Proceedings of the IEEE conference on computer vision and pattern recognition, (2018), 6848–6856. doi: 10.1109/CVPR.2018.00716. |
| [39] | F. Y. Shih, Image processing and pattern recognition: fundamentals and techniques, John Wiley & Sons, (2010). |
| [40] |
M. M. R. Krishnan, C. Chakraborty, R. R. Paul, A. K. Ray, Hybrid segmentation, characterization and classification of basal cell nuclei from histopathological images of normal oral mucosa and oral submucous fibrosis, Expert Syst. Appl., 39 (2012), 1062–1077. doi: 10.1016/j.eswa.2011.07.107. doi: 10.1016/j.eswa.2011.07.107
|
| [41] | L. Perez, J. Wang, The effectiveness of data augmentation in image classification using deep learning, (2017), arXiv preprint arXiv: 1712.04621. |
| [42] | F. Chollet, Xception: Deep learning with depthwise separable convolutions, in Proceedings of the IEEE conference on computer vision and pattern recognition, (2017), 1251–1258. doi: 10.1109/CVPR.2017.195 |
| [43] | K. He, X. Zhang, S. Ren, J. Sun, Deep residual learning for image recognition, in Proceedings of the IEEE conference on computer vision and pattern recognition, (2016). doi: 10.1109/CVPR.2016.90. |
| [44] | V. Nair, G. E. Hinton, Rectified linear units improve restricted Boltzmann machines, in International Conference on Machine Learning (ICML), (2010), 807–814. |
| [45] | W. Liu, Y. Wen, Z. Yu, M. Yang, Large-margin softmax loss for convolutional neural networks, in Proceedings of the 33rd International Conference on Machine Learning (2016), 507–516. |
| [46] | I. Sutskever, J. Martens, G. Dahl, G. Hinton, On the importance of initialization and momentum in deep learning, in International Conference on Machine Learning, PMLR, 28 (2013), 1139–1147. |
| [47] | D. P. Kingma, J. Ba, Adam: A method for stochastic optimization, (2014), arXiv preprint arXiv: 1412.6980. |
| [48] | K. He, J. Sun, Convolutional neural networks at constrained time cost, in Proceedings of the IEEE conference on computer vision and pattern recognition, (2015), 5353–5360. doi: 10.1109/CVPR.2015.7299173. |
| [49] |
T. Belvin, V. Kumar, S. Saini, Texture analysis based segmentation and classification of oral cancer lesions in color images using ANN, IEEE International Conference on Signal Processing, Computing and Control (ISPCC), (2013), 1–5. doi: 10.1109/ISPCC.2013.6663401. doi: 10.1109/ISPCC.2013.6663401
|
| [50] | K. Anuradha, K. Sankaranarayanan, Detection of Oral Tumors using Marker Controlled Segmentation, Int. J. Comp. Appl., 52 (2012), 15–18. |
| [51] |
B. R. Nanditha, A. Geetha, H. S. Chandrashekar, M. S. Dinesh, S. Murali, An Ensemble Deep Neural Network Approach for Oral Cancer Screening, Int. J. Online Biomed. Eng., 17 (2021), 121–134. doi: 10.3991/ijoe.v17i02.19207. doi: 10.3991/ijoe.v17i02.19207
|
Figures(7) / Tables(2)

Santisudha Panigrahi, Ruchi Bhuyan, Kundan Kumar, Janmenjoy Nayak, Tripti Swarnkar. Multistage classification of oral histopathological images using improved residual network[J]. Mathematical Biosciences and Engineering, 2022, 19(2): 1909-1925. doi: 10.3934/mbe.2022090







 DownLoad:
DownLoad: