Citation: Susanne Klawetter, Jennifer C. Greenfield, Stephanie Rachel Speer, Kyria Brown, Sunah S. Hwang. An integrative review: maternal engagement in the neonatal intensive care unit and health outcomes for U.S.-born preterm infants and their parents[J]. AIMS Public Health, 2019, 6(2): 160-183. doi: 10.3934/publichealth.2019.2.160

| [1] | Martin JA, Hamilton BE, Osterman MJ, et al. (2017) Births: Final Data for 2015. National vital statistics reports: from the Centers for Disease Control and Prevention, National Center for Health Statistics. Natl Vital Stat Rep 66: 1–2. |
| [2] |
Allen MC, Cristofalo EA, Kim C (2011) Outcomes of preterm infants: morbidity replaces mortality. Clin Perinatol 38: 441–454. doi: 10.1016/j.clp.2011.06.011

|
| [3] |
Cooke RJ (2016) Improving growth in preterm infants during initial hospital stay: principles into practice. Arch Dis Child Fetal Neonatal Ed 101: F366–370. doi: 10.1136/archdischild-2015-310097
|
| [4] |
Patel RM (2016) Short- and long-term outcomes for extremely preterm infants. Am J Perinatol 33: 318–328. doi: 10.1055/s-0035-1571202
|
| [5] | Maurer M, Dardess P, Carman KL, et al. (2013) Guide to patient and family engagement: environmental scan report. AHRQ Publication: No. 12-0042-EF. |
| [6] |
Samra HA, McGrath JM, Fischer S, et al. (2015) The NICU parent risk evaluation and engagement model and instrument (PREEMI) for neonates in intensive care units. J Obstet Gynecol Neonatal Nurs 44: 114–126. doi: 10.1111/1552-6909.12535
|
| [7] |
Richards CA, Starks H, O'Connor MR, et al. (2018) Physicians perceptions of shared decision-making in neonatal and pediatric critical care. Am J Hosp Palliat Care 35: 669–676. doi: 10.1177/1049909117734843
|
| [8] |
Celenza JF, Zayack D, Buus-Frank ME, et al. (2017) Family involvement in quality improvement: from bedside advocate to system advisor. Clin Perinatol 44: 553–566. doi: 10.1016/j.clp.2017.05.008
|
| [9] |
Davidson JE, Aslakson RA, Long AC, et al. (2017) Guidelines for family-centered care in the neonatal, pediatric, and adult ICU. Crit Care Med 45: 103–128. doi: 10.1097/CCM.0000000000002169
|
| [10] |
Weiss EM, Xie D, Cook N, et al. (2018) Characteristics associated with preferences for parent-centered decision making in neonatal intensive care. JAMA Pediatr 172: 461–468. doi: 10.1001/jamapediatrics.2017.5776
|
| [11] |
O'Brien K, Robson K, Bracht M, et al. (2018) Effectiveness of family integrated care in neonatal intensive care units on infant and parent outcomes: a multicentre, multinational, cluster-randomised controlled trial. Lancet Child Adolescent Health 2: 245–254. doi: 10.1016/S2352-4642(18)30039-7
|
| [12] | National Conference of State Legislatures: State and Family Medical Leave Laws, 2016. Available from: http://www.ncsl.org/research/labor-and-employment/state-family-and-medical-leave-laws.aspx. |
| [13] |
Hoffman C, Dunn DM, Njoroge WFM (2017) Impact of postpartum mental illness upon infant development. Curr Psychiatry Rep 19: 100. doi: 10.1007/s11920-017-0857-8
|
| [14] | Beck CT (1995) The effects of postpartum depression on maternal-infant interaction: a meta-analysis. Nurs Res 44: 298–304. |
| [15] |
Liberati A, Altman DG, Tetzlaff J, et al. (2009) The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. J Clin Epidemiol 62: e1–34. doi: 10.1016/j.jclinepi.2009.06.006
|
| [16] |
Neu M, Browne JV, Vojir C (2000) The impact of two transfer techniques used during skin-to-skin care on the physiologic and behavioral responses of preterm infants. Nurs Res 49: 215–23. doi: 10.1097/00006199-200007000-00005
|
| [17] |
Neu M (2014) An innovative investigation into the etiology of feeding intolerance in preterm infants. J Pediatr Gastroenterol Nutr 58: 4–5. doi: 10.1097/MPG.0000000000000188
|
| [18] | Ludington-Hoe SM, Anderson GC, Swinth JY, et al. (2004) Randomized controlled trial of kangaroo care: cardiorespiratory and thermal effects on healthy preterm infants. Neonatal Netw 23: 39–48. |
| [19] |
McCain GC, Ludington-Hoe SM, Swinth JY, et al. (2005) Heart rate variability responses of a preterm infant to kangaroo care. J Obstet Gynecol Neonatal Nurs 34: 689–694. doi: 10.1177/0884217505281857
|
| [20] |
Welch MG, Hofer MA, Stark RI, et al. (2013) Randomized controlled trial of family nurture intervention in the NICU: assessments of length of stay, feasibility and safety. BMC Pediatr 13: 148. doi: 10.1186/1471-2431-13-148
|
| [21] |
Cong X, Cusson RM, Walsh S, et al. (2012) Effects of skin-to-skin contact on autonomic pain responses in preterm infants. J Pain 13: 636–645. doi: 10.1016/j.jpain.2012.02.008
|
| [22] |
Cong X, Ludington-Hoe SM, McCain G, et al. (2009) Kangaroo care modifies preterm infant heart rate variability in response to heel stick pain: pilot study. Early Hum Dev 85: 561–567. doi: 10.1016/j.earlhumdev.2009.05.012
|
| [23] |
Reynolds LC, Duncan MM, Smith GC, et al. (2013) Parental presence and holding in the neonatal intensive care unit and associations with early neurobehavior. J Perinatol 33: 636–641. doi: 10.1038/jp.2013.4
|
| [24] |
Neu M, Robinson J, Schmiege SJ (2013) Influence of holding practice on preterm infant development. MCN Am J Matern Child Nurs 38: 136–143. doi: 10.1097/NMC.0b013e31827ca68c
|
| [25] |
Pineda R, Bender J, Hall B, et al. (2018) Parent participation in the neonatal intensive care unit: Predictors and relationships to neurobehavior and developmental outcomes. Early Hum Dev 117: 32–38. doi: 10.1016/j.earlhumdev.2017.12.008
|
| [26] |
Vittner D, McGrath J, Robinson J, et al. (2018) Increase in oxytocin from skin-to-skin contact enhances development of parent-infant relationship. Biol Res Nurs 20: 54–62. doi: 10.1177/1099800417735633
|
| [27] |
Zeskind PS, Iacino R (1984) Effects of maternal visitation to preterm infants in the neonatal intensive care unit. Child Dev 55: 1887–1893. doi: 10.2307/1129935
|
| [28] |
Whipple J (2000) The effect of parent training in music and multimodal stimulation on parent-neonate interactions in the neonatal intensive care unit. J Music Ther 37: 250–268. doi: 10.1093/jmt/37.4.250
|
| [29] | Lewis M, Bendersky M, Koons A, et al. (1991) Visitation to a neonatal intensive care unit. Pediatrics 88: 795–800. |
| [30] |
Greene MM, Rossman B, Patra K, et al. (2015) Maternal psychological distress and visitation to the neonatal intensive care unit. Acta Paediatr 104: e306–13. doi: 10.1111/apa.12975
|
| [31] |
Holditch-Davis D, White-Traut RC, Levy JA, et al. (2014) Maternally administered interventions for preterm infants in the NICU: effects on maternal psychological distress and mother-infant relationship. Infant Behav Dev 37: 695–710. doi: 10.1016/j.infbeh.2014.08.005
|
| [32] |
Lester BM, Salisbury AL, Hawes K, et al. (2016) 18-Month follow-up of infants cared for in a single-family room neonatal intensive care unit. J Pediatr 177: 84–89. doi: 10.1016/j.jpeds.2016.06.069
|
| [33] |
Gonya J, Ray WC, Rumpf RW, et al. (2017) Investigating skin-to-skin care patterns with extremely preterm infants in the NICU and their effect on early cognitive and communication performance: a retrospective cohort study. BMJ Open 7: e012985. doi: 10.1136/bmjopen-2016-012985
|
| [34] |
Welch MG, Firestein MR, Austin J, et al. (2015) Family nurture intervention in the neonatal intensive care unit improves social-relatedness, attention, and neurodevelopment of preterm infants at 18 months in a randomized controlled trial. J Child Psychol Psychiatry 56: 1202–1211. doi: 10.1111/jcpp.12405
|
| [35] |
Phillips RM, Merritt TA, Goldstein MR, et al. (2012) Prevention of postpartum smoking relapse in mothers of infants in the neonatal intensive care unit. J Perinatol 32: 374–380. doi: 10.1038/jp.2011.106
|
| [36] | Gale G, Franck L, Lund C (1993) Skin-to-skin (kangaroo) holding of the intubated premature infant. Neonatal Netw 12: 49–57. |
| [37] | Affonso D, Bosque E, Wahlberg V, et al. (1993) Reconciliation and healing for mothers through skin-to-skin contact provided in an American tertiary level intensive care nursery. Neonatal Netw 12: 25–32. |
| [38] |
Johnson AN (2007) The maternal experience of kangaroo holding. J Obstet Gynecol Neonatal Nurs 36: 568–573. doi: 10.1111/j.1552-6909.2007.00187.x
|
| [39] |
Roller CG (2005) Getting to know you: mothers' experiences of kangaroo care. J Obstet Gynecol Neonatal Nurs 34: 210–217. doi: 10.1177/0884217504273675
|
| [40] |
Cong X, Ludington-Hoe SM, Hussain N, et al. (2015) Parental oxytocin responses during skin-to-skin contact in pre-term infants. Early Hum Dev 91: 401–406. doi: 10.1016/j.earlhumdev.2015.04.012
|
| [41] |
Samra HA, Dutcher J, McGrath JM, et al. (2015) Effect of skin-to-skin holding on stress in mothers of late-preterm infants: A randomized controlled trial. Adv Neonatal Care 15: 354–364. doi: 10.1097/ANC.0000000000000223
|
| [42] |
Bloch-Salisbury E, Zuzarte I, Indic P, et al. (2014) Kangaroo care: cardio-respiratory relationships between the infant and caregiver. Early Hum Dev 90: 843–850. doi: 10.1016/j.earlhumdev.2014.08.015
|
| [43] | Neu M, Laudenslager ML, Robinson J (2009) Coregulation in salivary cortisol during maternal holding of premature infants. Biol Res Nurs 10: 226–240. |
| [44] |
Neu M, Hazel NA, Robinson J, et al. (2014) Effect of holding on co-regulation in preterm infants: a randomized controlled trial. Early Hum Dev 90: 141–147. doi: 10.1016/j.earlhumdev.2014.01.008
|
| [45] |
Hane AA, Myers MM, Hofer MA, et al. (2015) Family nurture intervention improves the quality of maternal caregiving in the neonatal intensive care unit: evidence from a randomized controlled trial. J Dev Behav Pediatr 36: 188–196. doi: 10.1097/DBP.0000000000000148
|
| [46] |
White-Traut R, Wink T, Minehart T, et al. (2012) Frequency of premature infant engagement and disengagement behaviors during two maternally administered interventions. Newborn Infant Nurs Rev 12: 124–131. doi: 10.1053/j.nainr.2012.06.005
|
| [47] | Hurst NM, Valentine CJ, Renfro L, et al. (1997) Skin-to-skin holding in the neonatal intensive care unit influences maternal milk volume. J Perinatol 17: 213–217. |
| [48] |
Lau C, Hurst NM, Smith EO, et al. (2007) Ethnic/racial diversity, maternal stress, lactation and very low birthweight infants. J Perinatol 27: 399–408. doi: 10.1038/sj.jp.7211770
|
| [49] |
Tully KP, Holditch-Davis D, White-Traut RC, et al. (2016) A test of kangaroo care on preterm infant breastfeeding. J Obstet Gynecol Neonatal Nurs 45: 45–61. doi: 10.1016/j.jogn.2015.10.004
|
| [50] |
Dieterich CM, Felice JP, O'Sullivan E, et al. (2013) Breastfeeding and health outcomes for the mother-infant dyad. Pediatr Clin North Am 60: 31–48. doi: 10.1016/j.pcl.2012.09.010
|
| [51] | Li R, Dee D, Li CM, et al. (2014) Breastfeeding and risk of infections at 6 years. Pediatrics 134 Suppl 1: S13–20. |
| [52] | Thompson JMD, Tanabe K, Moon RY, et al. (2017) Duration of breastfeeding and risk of SIDS: An individual participant data meta-analysis. Pediatrics 140. |
| [53] |
Salihu HM, Wilson RE (2007) Epidemiology of prenatal smoking and perinatal outcomes. Early Hum Dev 83: 713–720. doi: 10.1016/j.earlhumdev.2007.08.002
|
| [54] |
Herrmann M, King K, Weitzman M (2008) Prenatal tobacco smoke and postnatal secondhand smoke exposure and child neurodevelopment. Curr Opin Pediatr 20: 184–190. doi: 10.1097/MOP.0b013e3282f56165
|
| [55] |
Zhou S, Rosenthal DG, Sherman S, et al. (2014) Physical, behavioral, and cognitive effects of prenatal tobacco and postnatal secondhand smoke exposure. Curr Probl Pediatr Adolesc Health Care 44: 219–241. doi: 10.1016/j.cppeds.2014.03.007
|
| [56] | Dunn MS, Reilly MC, Johnston AM, et al. (2006) Development and dissemination of potentially better practices for the provision of family-centered care in neonatology: the family-centered care map. Pediatrics 118 Suppl 2: S95–107. |
| [57] |
Stefana A, Lavelli M (2017) Parental engagement and early interactions with preterm infants during the stay in the neonatal intensive care unit: protocol of a mixed-method and longitudinal study. BMJ Open 7: e013824. doi: 10.1136/bmjopen-2016-013824
|
| [58] | O'Brien K, Bracht M, Macdonell K, et al. (2013) A pilot cohort analytic study of family integrated care in a Canadian neonatal intensive care unit. BMC Pregnancy Childbirth 13 Suppl 1: S12. |
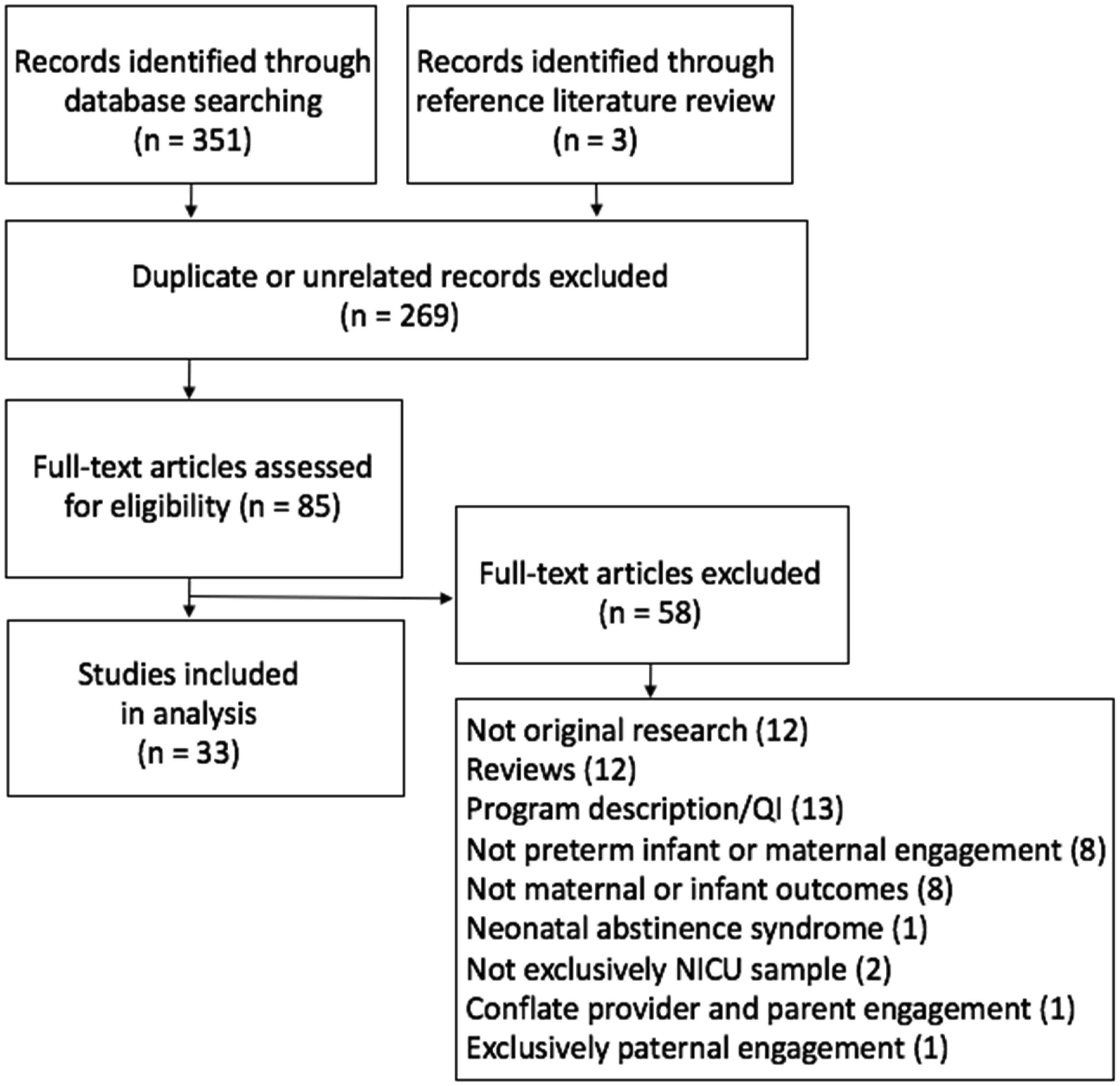
Figures(1) / Tables(1)

Susanne Klawetter, Jennifer C. Greenfield, Stephanie Rachel Speer, Kyria Brown, Sunah S. Hwang. An integrative review: maternal engagement in the neonatal intensive care unit and health outcomes for U.S.-born preterm infants and their parents[J]. AIMS Public Health, 2019, 6(2): 160-183. doi: 10.3934/publichealth.2019.2.160







 DownLoad:
DownLoad: