Citation: Mi Young Son, Paul Hasty. Homologous recombination defects and how they affect replication fork maintenance[J]. AIMS Genetics, 2018, 5(4): 192-211. doi: 10.3934/genet.2018.4.192

| [1] |
Bhargava R, Onyango DO, Stark JM (2016) Regulation of single-strand annealing and its role in genome maintenance. Trends Genet 32: 566–575. doi: 10.1016/j.tig.2016.06.007

|
| [2] |
Mizuno K, Lambert S, Baldacci G, et al. (2009) Nearby inverted repeats fuse to generate acentric and dicentric palindromic chromosomes by a replication template exchange mechanism. Genes Dev 23: 2876–2886. doi: 10.1101/gad.1863009
|
| [3] |
Petermann E, Orta ML, Issaeva N, et al. (2010) Hydroxyurea-stalled replication forks become progressively inactivated and require two different RAD51-mediated pathways for restart and repair. Mol Cell 37: 492–502. doi: 10.1016/j.molcel.2010.01.021
|
| [4] |
Kim TM, Ko JH, Hu L (2012) RAD51 mutants cause replication defects and chromosomal instability. Mol Cell Biol 32: 3663–3680. doi: 10.1128/MCB.00406-12
|
| [5] |
Schlacher K, Wu H, Jasin M (2012) A distinct replication fork protection pathway connects fanconi anemia tumor suppressors to RAD51-BRCA1/2. Cancer Cell 22: 106–116. doi: 10.1016/j.ccr.2012.05.015
|
| [6] |
Carr AM, Lambert S (2013) Replication stress-induced genome instability: The dark side of replication maintenance by homologous recombination. J Mol Biol 425: 4733–4744. doi: 10.1016/j.jmb.2013.04.023
|
| [7] |
Krejci L, Altmannova V, Spirek M, et al. (2012) Homologous recombination and its regulation. Nucleic Acids Res 40: 5795–5818. doi: 10.1093/nar/gks270
|
| [8] |
Wu Y, Kantake N, Sugiyama T, et al. (2008) Rad51 protein controls Rad52-mediated DNA annealing. J Biol Chem 283: 14883–14892. doi: 10.1074/jbc.M801097200
|
| [9] |
Yu DS, Sonoda E, Takeda S, et al. (2003) Dynamic control of Rad51 recombinase by self-association and interaction with BRCA2. Mol Cell 12: 1029–1041. doi: 10.1016/S1097-2765(03)00394-0
|
| [10] |
Ma CJ, Gibb B, Kwon Y, et al. (2017) Protein dynamics of human RPA and RAD51 on ssDNA during assembly and disassembly of the RAD51 filament. Nucleic Acids Res 45: 749–761. doi: 10.1093/nar/gkw1125
|
| [11] |
Chen R, Wold MS (2014) Replication protein A: Single-stranded DNA's first responder: Dynamic DNA-interactions allow replication protein A to direct single-strand DNA intermediates into different pathways for synthesis or repair. Bioessays 36: 1156–1161. doi: 10.1002/bies.201400107
|
| [12] |
Ma CJ, Kwon Y, Sung P, et al. (2017) Human RAD52 interactions with replication protein A and the RAD51 presynaptic complex. J Biol Chem 292: 11702–11713. doi: 10.1074/jbc.M117.794545
|
| [13] |
van der Heijden T, Seidel R, Modesti M, et al. (2007) Real-time assembly and disassembly of human RAD51 filaments on individual DNA molecules. Nucleic Acids Res 35: 5646–5657. doi: 10.1093/nar/gkm629
|
| [14] |
Candelli A, Holthausen JT, Depken M, et al. (2014) Visualization and quantification of nascent RAD51 filament formation at single-monomer resolution. Proc Natl Acad Sci USA 111: 15090–15095. doi: 10.1073/pnas.1307824111
|
| [15] |
Prakash R, Zhang Y, Feng W, et al. (2015) Homologous recombination and human health: The roles of BRCA1, BRCA2, and associated proteins. Cold Spring Harb Perspect Biol 7: a016600. doi: 10.1101/cshperspect.a016600
|
| [16] |
Oh J, Symington LS (2018) Role of the Mre11 complex in preserving genome integrity. Genes 9: 589. doi: 10.3390/genes9120589
|
| [17] |
Yu X, Wu LC, Bowcock AM, et al. (1998) The C-terminal (BRCT) domains of BRCA1 interact in vivo with CtIP, a protein implicated in the CtBP pathway of transcriptional repression. J Biol Chem 273: 25388–25392. doi: 10.1074/jbc.273.39.25388
|
| [18] |
Zhong Q, Chen CF, Li S, et al. (1999) Association of BRCA1 with the hRad50-hMre11-p95 complex and the DNA damage response. Science 285: 747–750. doi: 10.1126/science.285.5428.747
|
| [19] |
Xia B, Sheng Q, Nakanishi K, et al. (2006) Control of BRCA2 cellular and clinical functions by a nuclear partner, PALB2. Mol Cell 22: 719–729. doi: 10.1016/j.molcel.2006.05.022
|
| [20] |
Sy SM, Huen MS, Chen J (2009) PALB2 is an integral component of the BRCA complex required for homologous recombination repair. Proc Natl Acad Sci USA 106: 7155–7160. doi: 10.1073/pnas.0811159106
|
| [21] | Zhang F, Ma J, Wu J (2009) PALB2 links BRCA1 and BRCA2 in the DNA-damage response. Curr Biol 19: 524–529. |
| [22] |
Zhang F, Fan Q, Ren K, et al. (2009) PALB2 functionally connects the breast cancer susceptibility proteins BRCA1 and BRCA2. Mol Cancer Res 7: 1110–1118. doi: 10.1158/1541-7786.MCR-09-0123
|
| [23] |
Yang H, Li Q, Fan J, et al. (2005) The BRCA2 homologue Brh2 nucleates RAD51 filament formation at a dsDNA-ssDNA junction. Nature 433: 653–657. doi: 10.1038/nature03234
|
| [24] |
Jensen RB, Carreira A, Kowalczykowski SC (2010) Purified human BRCA2 stimulates RAD51-mediated recombination. Nature 467: 678–683. doi: 10.1038/nature09399
|
| [25] |
Liu J, Doty T, Gibson B, et al. (2010) Human BRCA2 protein promotes RAD51 filament formation on RPA-covered single-stranded DNA. Nat Struct Mol Biol 17: 1260–1262. doi: 10.1038/nsmb.1904
|
| [26] |
Thorslund T, McIlwraith MJ, Compton SA, et al. (2010) The breast cancer tumor suppressor BRCA2 promotes the specific targeting of RAD51 to single-stranded DNA. Nat Struct Mol Biol 17: 1263–1265. doi: 10.1038/nsmb.1905
|
| [27] |
Quinet A, Lemacon D, Vindigni A (2017) Replication Fork Reversal: Players and Guardians. Mol Cell 68: 830–833. doi: 10.1016/j.molcel.2017.11.022
|
| [28] |
San Filippo J, Chi P, Sehorn MG, et al. (2006) Recombination mediator and Rad51 targeting activities of a human BRCA2 polypeptide. J Biol Chem 281: 11649–11657. doi: 10.1074/jbc.M601249200
|
| [29] |
Chen P, Chen C, Chen Y, et al. (1998) The BRC repeats in BRCA2 are critical for RAD51 binding and resistance to methyl methanesulfonate treatment. Proc Natl Acad Sci USA 95: 5287–5292. doi: 10.1073/pnas.95.9.5287
|
| [30] |
Chen C, Chen P, Zhong Q, et al. (1999) Expression of BRC repeats in breast cancer cells disrupts the BRCA2-Rad51 complex and leads to radiation hypersensitivity and loss of G(2)/M checkpoint control. J Biol Chem 274: 32931–32935. doi: 10.1074/jbc.274.46.32931
|
| [31] |
Rajendra E, Venkitaraman AR (2010) Two modules in the BRC repeats of BRCA2 mediate structural and functional interactions with the RAD51 recombinase. Nucleic Acids Res 38: 82–96. doi: 10.1093/nar/gkp873
|
| [32] |
Carreira A, Kowalczykowski SC (2011) Two classes of BRC repeats in BRCA2 promote RAD51 nucleoprotein filament function by distinct mechanisms. Proc Natl Acad Sci USA 108: 10448–10453. doi: 10.1073/pnas.1106971108
|
| [33] |
Galkin VE, Esashi F, Yu X, et al. (2005) BRCA2 BRC motifs bind RAD51-DNA filaments. Proc Natl Acad Sci USA 102: 8537–8542. doi: 10.1073/pnas.0407266102
|
| [34] |
Carreira A, Hilario J, Amitani I, et al. (2009) The BRC repeats of BRCA2 modulate the DNA-binding selectivity of RAD51. Cell 136: 1032–1043. doi: 10.1016/j.cell.2009.02.019
|
| [35] |
Chatterjee G, Jimenez-Sainz J, Presti T, et al. (2016) Distinct binding of BRCA2 BRC repeats to RAD51 generates differential DNA damage sensitivity. Nucleic Acids Res 44: 5256–5270. doi: 10.1093/nar/gkw242
|
| [36] |
Sharan SK, Morimatsu M, Albrecht U, et al. (1997) Embryonic lethality and radiation hypersensitivity mediated by Rad51 in mice lacking Brca2. Nature 386: 804–810. doi: 10.1038/386804a0
|
| [37] | Morimatsu M, Donoho G, Hasty P (1998) Cells deleted for Brca2 COOH terminus exhibit hypersensitivity to gamma-radiation and premature senescence. Cancer Res 58: 3441–3447. |
| [38] |
Davies OR, Pellegrini L (2007) Interaction with the BRCA2 C terminus protects RAD51-DNA filaments from disassembly by BRC repeats. Nat Struct Mol Biol 14: 475–483. doi: 10.1038/nsmb1251
|
| [39] |
Esashi F, Galkin VE, Yu X, et al. (2007) Stabilization of RAD51 nucleoprotein filaments by the C-terminal region of BRCA2. Nat Struct Mol Biol 14: 468–474. doi: 10.1038/nsmb1245
|
| [40] |
Schlacher K, Christ N, Siaud N, et al. (2011) Double-Strand Break Repair-Independent Role for BRCA2 in Blocking Stalled Replication Fork Degradation by MRE11. Cell 145: 529–542. doi: 10.1016/j.cell.2011.03.041
|
| [41] |
Ayoub N, Rajendra E, Su X, et al. (2009) The carboxyl terminus of Brca2 links the disassembly of Rad51 complexes to mitotic entry. Curr Biol 19: 1075–1085. doi: 10.1016/j.cub.2009.05.057
|
| [42] | Kim TM, Son MY, Dodds S, et al. (2014) Deletion of BRCA2 exon 27 causes defects in response to both stalled and collapsed replication forks. Mutat Res 766–767: 66–72. |
| [43] |
Donoho G, Brenneman MA, Cui TX, et al. (2003) Deletion of Brca2 exon 27 causes hypersensitivity to DNA crosslinks, chromosomal instability, and reduced life span in mice. Genes Chromosomes Cancer 36: 317–331. doi: 10.1002/gcc.10148
|
| [44] |
Taylor MRG, Spirek M, Chaurasiya KR, et al. (2015) Rad51 Paralogs Remodel Pre-synaptic Rad51 Filaments to Stimulate Homologous Recombination. Cell 162: 271–286. doi: 10.1016/j.cell.2015.06.015
|
| [45] |
Taylor MRG, Spirek M, Jian Ma C, et al. (2016) A Polar and Nucleotide-Dependent Mechanism of Action for RAD51 Paralogs in RAD51 Filament Remodeling. Mol Cell 64: 926–939. doi: 10.1016/j.molcel.2016.10.020
|
| [46] |
Masson JY, Tarsounas MC, Stasiak AZ, et al. (2001) Identification and purification of two distinct complexes containing the five RAD51 paralogs. Genes Dev 15: 3296–3307. doi: 10.1101/gad.947001
|
| [47] |
Chun J, Buechelmaier ES, Powell SN (2013) Rad51 paralog complexes BCDX2 and CX3 act at different stages in the BRCA1-BRCA2-dependent homologous recombination pathway. Mol Cell Biol 33: 387–395. doi: 10.1128/MCB.00465-12
|
| [48] |
Sigurdsson S, Van Komen S, Bussen W, et al. (2001) Mediator function of the human Rad51B-Rad51C complex in Rad51/RPA-catalyzed DNA strand exchange. Genes Dev 15: 3308–3318. doi: 10.1101/gad.935501
|
| [49] |
Kurumizaka H, Ikawa S, Nakada M, et al. (2001) Homologous-pairing activity of the human DNA-repair proteins Xrcc3.Rad51C. Proc Natl Acad Sci USA 98: 5538–5543. doi: 10.1073/pnas.091603098
|
| [50] |
Kurumizaka H, Ikawa S, Nakada M, et al. (2002) Homologous pairing and ring and filament structure formation activities of the human Xrcc2*Rad51D complex. J Biol Chem 277: 14315–14320. doi: 10.1074/jbc.M105719200
|
| [51] |
Yard BD, Reilly NM, Bedenbaugh MK, et al. (2016) RNF138 interacts with RAD51D and is required for DNA interstrand crosslink repair and maintaining chromosome integrity. DNA Repair 42: 82–93. doi: 10.1016/j.dnarep.2016.04.006
|
| [52] |
Piwko W, Mlejnkova LJ, Mutreja K, et al. (2016) The MMS22L-TONSL heterodimer directly promotes RAD51-dependent recombination upon replication stress. EMBO J 35: 2584–2601. doi: 10.15252/embj.201593132
|
| [53] |
Huang TH, Fowler F, Chen CC, et al. (2018) The histone chaperones ASF1 and CAF-1 promote MMS22L-TONSL-Mediated RAD51 loading onto ssDNA during homologous recombination in human cells. Mol Cell 69: 879–892. doi: 10.1016/j.molcel.2018.01.031
|
| [54] |
O'Donnell L, Panier S, Wildenhain J, et al. (2010) The MMS22L-TONSL complex mediates recovery from replication stress and homologous recombination. Mol Cell 40: 619–631. doi: 10.1016/j.molcel.2010.10.024
|
| [55] |
Duro E, Lundin C, Ask K, et al. (2010) Identification of the MMS22L-TONSL complex that promotes homologous recombination. Mol Cell 40: 632–644. doi: 10.1016/j.molcel.2010.10.023
|
| [56] |
Higgs MR, Reynolds JJ, Winczura A, et al. (2015) BOD1L is required to suppress deleterious resection of stressed replication forks. Mol Cell 59: 462–477. doi: 10.1016/j.molcel.2015.06.007
|
| [57] |
Modesti M, Budzowska M, Baldeyron C, et al. (2007) RAD51AP1 is a structure-specific DNA binding protein that stimulates joint molecule formation during RAD51-mediated homologous recombination. Mol Cell 28: 468–481. doi: 10.1016/j.molcel.2007.08.025
|
| [58] |
Wiese C, Dray E, Groesser T, et al. (2007) Promotion of homologous recombination and genomic stability by RAD51AP1 via RAD51 recombinase enhancement. Mol Cell 28: 482–490. doi: 10.1016/j.molcel.2007.08.027
|
| [59] |
Lu CH, Yeh HY, Su GC, et al. (2018) Swi5-Sfr1 stimulates Rad51 recombinase filament assembly by modulating Rad51 dissociation. Proc Natl Acad Sci USA 115: E10059–E10068. doi: 10.1073/pnas.1812753115
|
| [60] |
Forget AL, Loftus MS, McGrew DA, et al. (2007) The human Rad51 K133A mutant is functional for DNA double-strand break repair in human cells. Biochemistry 46: 3566–3575. doi: 10.1021/bi062128k
|
| [61] |
Chi P, Van Komen S, Sehorn MG, et al. (2006) Roles of ATP binding and ATP hydrolysis in human Rad51 recombinase function. DNA Repair 5: 381–391. doi: 10.1016/j.dnarep.2005.11.005
|
| [62] |
Stark JM, Hu P, Pierce AJ, et al. (2002) ATP hydrolysis by mammalian RAD51 has a key role during homology-directed DNA repair. J Biol Chem 277: 20185–20194. doi: 10.1074/jbc.M112132200
|
| [63] |
Brouwer I, Moschetti T, Candelli A, et al. (2018) Two distinct conformational states define the interaction of human RAD51-ATP with single-stranded DNA. EMBO J 37: e98162. doi: 10.15252/embj.201798162
|
| [64] |
van Mameren J, Modesti M, Kanaar R, et al. (2009) Counting RAD51 proteins disassembling from nucleoprotein filaments under tension. Nature 457: 745–748. doi: 10.1038/nature07581
|
| [65] |
Yu X, Jacobs SA, West SC, et al. (2001) Domain structure and dynamics in the helical filaments formed by RecA and Rad51 on DNA. Proc Natl Acad Sci USA 98: 8419–8424. doi: 10.1073/pnas.111005398
|
| [66] | Short JM, Liu Y, Chen S, et al. (2016) High-resolution structure of the presynaptic RAD51 filament on single-stranded DNA by electron cryo-microscopy. Nucleic Acids Res 44: 9017–9030. |
| [67] |
Spirek M, Mlcouskova J, Belan O, et al. (2018) Human RAD51 rapidly forms intrinsically dynamic nucleoprotein filaments modulated by nucleotide binding state. Nucleic Acids Res 46: 3967–3980. doi: 10.1093/nar/gky111
|
| [68] |
Kelso AA, Goodson SD, Watts LE, et al. (2017) The beta-isoform of BCCIP promotes ADP release from the RAD51 presynaptic filament and enhances homologous DNA pairing. Nucleic Acids Res 45: 711–725. doi: 10.1093/nar/gkw877
|
| [69] |
Popova M, Shimizu H, Yamamoto K, et al. (2009) Detection of c-Abl kinase-promoted phosphorylation of Rad51 by specific antibodies reveals that Y54 phosphorylation is dependent on that of Y315. FEBS Lett 583: 1867–1872. doi: 10.1016/j.febslet.2009.04.044
|
| [70] |
Pakakasama S, Kajanachumpol S, Kanjanapongkul S, et al. (2008) Simple multiplex RT-PCR for identifying common fusion transcripts in childhood acute leukemia. Int J Lab Hematol 30: 286–291. doi: 10.1111/j.1751-553X.2007.00954.x
|
| [71] | Mencalha AL, Correa S, Abdelhay E (2014) Role of calcium-dependent protein kinases in chronic myeloid leukemia: Combined effects of PKC and BCR-ABL signaling on cellular alterations during leukemia development. Onco Targets Ther 7: 1247–1254. |
| [72] |
Maiani E, Diederich M, Gonfloni S (2011) DNA damage response: The emerging role of c-Abl as a regulatory switch? Biochem Pharmacol 82: 1269–1276. doi: 10.1016/j.bcp.2011.07.001
|
| [73] |
Slupianek A, Dasgupta Y, Ren SY, et al. (2011) Targeting RAD51 phosphotyrosine-315 to prevent unfaithful recombination repair in BCR-ABL1 leukemia. Blood 118: 1062–1068. doi: 10.1182/blood-2010-09-307256
|
| [74] |
Shimizu H, Popova M, Fleury F, et al. (2009) c-ABL tyrosine kinase stabilizes RAD51 chromatin association. Biochem Biophys Res Commun 382: 286–291. doi: 10.1016/j.bbrc.2009.03.020
|
| [75] |
Yuan ZM, Huang Y, Ishiko T, et al. (1998) Regulation of Rad51 function by c-Abl in response to DNA damage. J Biol Chem 273: 3799–3802. doi: 10.1074/jbc.273.7.3799
|
| [76] |
Subramanyam S, Ismail M, Bhattacharya I, et al. (2016) Tyrosine phosphorylation stimulates activity of human RAD51 recombinase through altered nucleoprotein filament dynamics. Proc Natl Acad Sci USA 113: E6045–E6054. doi: 10.1073/pnas.1604807113
|
| [77] |
Alligand B, Le Breton M, Marquis D, et al. (2017) Functional effects of diphosphomimetic mutations at cAbl-mediated phosphorylation sites on Rad51 recombinase activity. Biochimie 139: 115–124. doi: 10.1016/j.biochi.2017.05.020
|
| [78] |
Sorensen CS, Hansen LT, Dziegielewski J, et al. (2005) The cell-cycle checkpoint kinase Chk1 is required for mammalian homologous recombination repair. Nat Cell Biol 7: 195–201. doi: 10.1038/ncb1212
|
| [79] |
Yata K, Lloyd J, Maslen S, et al. (2012) Plk1 and CK2 act in concert to regulate Rad51 during DNA double strand break repair. Mol Cell 45:371–383. doi: 10.1016/j.molcel.2011.12.028
|
| [80] |
Hu L, Kim TM, Son MY, et al. (2013) Two replication fork maintenance pathways fuse inverted repeats to rearrange chromosomes. Nature 501: 569–572. doi: 10.1038/nature12500
|
| [81] |
Chapman JR, Taylor MR, Boulton SJ (2012) Playing the end game: DNA double-strand break repair pathway choice. Mol Cell 47: 497–510. doi: 10.1016/j.molcel.2012.07.029
|
| [82] |
Hu Y, Raynard S, Sehorn MG, et al. (2007) RECQL5/Recql5 helicase regulates homologous recombination and suppresses tumor formation via disruption of Rad51 presynaptic filaments. Genes Dev 21: 3073–3084. doi: 10.1101/gad.1609107
|
| [83] |
Sommers JA, Rawtani N, Gupta R, et al. (2009) FANCJ uses its motor ATPase to destabilize protein-DNA complexes, unwind triplexes, and inhibit RAD51 strand exchange. J Biol Chem 284: 7505–7517. doi: 10.1074/jbc.M809019200
|
| [84] |
Di Marco S, Hasanova Z, Kanagaraj R, et al. (2017) RECQ5 Helicase Cooperates with MUS81 Endonuclease in Processing Stalled Replication Forks at Common Fragile Sites during Mitosis. Mol Cell 66: 658–671. doi: 10.1016/j.molcel.2017.05.006
|
| [85] |
Bugreev DV, Yu X, Egelman EH, et al. (2007) Novel pro- and anti-recombination activities of the Bloom's syndrome helicase. Genes Dev 21: 3085–3094. doi: 10.1101/gad.1609007
|
| [86] |
Wu L, Hickson ID (2003) The Bloom's syndrome helicase suppresses crossing over during homologous recombination. Nature 426: 870–874. doi: 10.1038/nature02253
|
| [87] |
Gari K, Decaillet C, Stasiak AZ, et al. (2008) The Fanconi anemia protein FANCM can promote branch migration of Holliday junctions and replication forks. Mol Cell 29: 141–148. doi: 10.1016/j.molcel.2007.11.032
|
| [88] |
Gali H, Juhasz S, Morocz M, et al. (2012) Role of SUMO modification of human PCNA at stalled replication fork. Nucleic Acids Res 40: 6049–6059. doi: 10.1093/nar/gks256
|
| [89] |
Moldovan GL, Dejsuphong D, Petalcorin MI, et al. (2012) Inhibition of homologous recombination by the PCNA-interacting protein PARI. Mol Cell 45: 75–86. doi: 10.1016/j.molcel.2011.11.010
|
| [90] |
Burkovics P, Dome L, Juhasz S, et al. (2016) The PCNA-associated protein PARI negatively regulates homologous recombination via the inhibition of DNA repair synthesis. Nucleic Acids Res 44: 3176–3189. doi: 10.1093/nar/gkw024
|
| [91] |
Meyerson M, Pellman D (2011) Cancer genomes evolve by pulverizing single chromosomes. Cell 144: 9–10. doi: 10.1016/j.cell.2010.12.025
|
| [92] |
Scully R, Chen J, Plug A, et al. (1997) Association of BRCA1 with Rad51 in mitotic and meiotic cells. Cell 88: 265–275. doi: 10.1016/S0092-8674(00)81847-4
|
| [93] | Chen JJ, Silver D, Cantor S, et al. (1999) BRCA1, BRCA2, and Rad51 operate in a common DNA damage response pathway. Cancer Res 59: 1752s–1756s. |
| [94] | BCLC (1999) Cancer risks in BRCA2 mutation carriers. The breast cancer linkage consortium. J Natl Cancer Inst 91: 1310–1316. |
| [95] |
Jasin M (2002) Homologous repair of DNA damage and tumorigenesis: The BRCA connection. Oncogene 21: 8981–8993. doi: 10.1038/sj.onc.1206176
|
| [96] |
Ibrahim M, Yadav S, Ogunleye F, et al. (2018) Male BRCA mutation carriers: Clinical characteristics and cancer spectrum. BMC Cancer 18: 179. doi: 10.1186/s12885-018-4098-y
|
| [97] |
Lim DS, Hasty P (1996) A mutation in mouse rad51 results in an early embryonic lethal that is suppressed by a mutation in p53. Mol Cell Biol 16: 7133–7143. doi: 10.1128/MCB.16.12.7133
|
| [98] | Ceccaldi R, Sarangi P, D'Andrea AD (2016) The Fanconi anaemia pathway: new players and new functions. Nat Rev Mol Cell Biol 17: 337–349. |
| [99] |
Zadorozhny K, Sannino V, Belan O, et al. (2017) Fanconi-Anemia-Associated mutations destabilize RAD51 filaments and impair replication fork protection. Cell Rep 21: 333–340. doi: 10.1016/j.celrep.2017.09.062
|
| [100] |
Wang AT, Kim T, Wagner JE, et al. (2015) A dominant mutation in human RAD51 reveals its function in DNA interstrand crosslink repair independent of homologous recombination. Mol Cell 59: 478–490. doi: 10.1016/j.molcel.2015.07.009
|
| [101] |
Yang X, Wang A (1999) Structural studies of atom-specific anticancer drugs acting on DNA. Pharmacol Ther 83: 181–215. doi: 10.1016/S0163-7258(99)00020-0
|
| [102] |
Bessho T (2003) Induction of DNA replication-mediated double strand breaks by psoralen DNA interstrand cross-links. J Biol Chem 278: 5250–5254. doi: 10.1074/jbc.M212323200
|
| [103] |
Ameziane N, May P, Haitjema A, et al. (2015) A novel Fanconi anaemia subtype associated with a dominant-negative mutation in RAD51. Nat Commun 6: 8829. doi: 10.1038/ncomms9829
|
| [104] |
Silva MC, Morrical MD, Bryan KE, et al. (2016) RAD51 variant proteins from human lung and kidney tumors exhibit DNA strand exchange defects. DNA Repair 42: 44–55. doi: 10.1016/j.dnarep.2016.02.008
|
| [105] |
Lose F, Lovelock P, Chenevix-Trench G, et al. (2006) Variation in the RAD51 gene and familial breast cancer. Breast Cancer Res 8: R26. doi: 10.1186/bcr1415
|
| [106] |
Sun M, Ju H, Zhou Zh, et al. (2015) Genetic variants of the DNA damage repair genes XRCC4 and RAD51 are associated with susceptibility to esophageal cancer. Clin Res Hepatol Gastroenterol 39: 379–383. doi: 10.1016/j.clinre.2014.12.002
|
| [107] |
Zhao M, Chen P, Dong Y, et al. (2014) Relationship between Rad51 G135C and G172T variants and the susceptibility to cancer: A meta-analysis involving 54 case-control studies. PLoS One 9: e87259. doi: 10.1371/journal.pone.0087259
|
| [108] |
Klein HL (2008) The consequences of Rad51 overexpression for normal and tumor cells. DNA Repair 7: 686–693. doi: 10.1016/j.dnarep.2007.12.008
|
| [109] |
Depienne C, Bouteiller D, Meneret A, et al. (2012) RAD51 haploinsufficiency causes congenital mirror movements in humans. Am J Hum Genet 90: 301–307. doi: 10.1016/j.ajhg.2011.12.002
|
| [110] |
Gildemeister OS, Sage JM, Knight KL (2009) Cellular redistribution of Rad51 in response to DNA damage: Novel role for Rad51C. J Biol Chem 284: 31945–31952. doi: 10.1074/jbc.M109.024646
|
| [111] |
Jeyasekharan AD, Liu Y, Hattori H, et al. (2013) A cancer-associated BRCA2 mutation reveals masked nuclear export signals controlling localization. Nat Struct Mol Biol 20: 1191–1198. doi: 10.1038/nsmb.2666
|
| [112] |
Kutay U, Bischoff FR, Kostka S, et al. (1997) Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell 90: 1061–1071. doi: 10.1016/S0092-8674(00)80372-4
|
| [113] |
Okimoto S, Sun J, Fukuto A, et al. (2015) hCAS/CSE1L regulates RAD51 distribution and focus formation for homologous recombinational repair. Genes Cells 20: 681–694. doi: 10.1111/gtc.12262
|
| [114] |
Alshareeda AT, Negm OH, Green AR, et al. (2015) KPNA2 is a nuclear export protein that contributes to aberrant localisation of key proteins and poor prognosis of breast cancer. Br J Cancer 112: 1929–1937. doi: 10.1038/bjc.2015.165
|
| [115] |
Lee YJ, Sheu TJ, Keng PC (2005) Enhancement of radiosensitivity in H1299 cancer cells by actin-associated protein cofilin. Biochem Biophys Res Commun 335: 286–291. doi: 10.1016/j.bbrc.2005.07.073
|
| [116] | Hasty P, Christy BA (2013) p53 as an intervention target for cancer and aging. Pathobiol Aging Age Relat Dis 3. |
| [117] |
Gatz SA, Wiesmuller L (2006) p53 in recombination and repair. Cell Death Differ 13: 1003–1016. doi: 10.1038/sj.cdd.4401903
|
| [118] |
Janz C, Wiesmuller L (2002) Wild-type p53 inhibits replication-associated homologous recombination. Oncogene 21: 5929–5933. doi: 10.1038/sj.onc.1205757
|
| [119] | Linke SP, Sengupta S, Khabie N, et al. (2003) p53 interacts with hRAD51 and hRAD54, and directly modulates homologous recombination. Cancer Res 63: 2596–2605. |
| [120] |
Sturzbecher HW, Donzelmann B, Henning W, et al. (1996) p53 is linked directly to homologous recombination processes via RAD51/RecA protein interaction. EMBO J 15: 1992–2002. doi: 10.1002/j.1460-2075.1996.tb00550.x
|
| [121] |
Janz C, Susse S, Wiesmuller L (2002) p53 and recombination intermediates: Role of tetramerization at DNA junctions in complex formation and exonucleolytic degradation. Oncogene 21: 2130–2140. doi: 10.1038/sj.onc.1205292
|
| [122] |
Yoon D, Wang Y, Stapleford K, et al. (2004) p53 inhibits strand exchange and replication fork regression promoted by human Rad51. J Mol Biol 336: 639–654. doi: 10.1016/j.jmb.2003.12.050
|
| [123] |
Buchhop S, Gibson MK, Wang XW, et al. (1997) Interaction of p53 with the human Rad51 protein. Nucleic Acids Res 25: 3868–3874. doi: 10.1093/nar/25.19.3868
|
| [124] |
Duan W, Ding H, Subler MA, et al. (2002) Lung-specific expression of human mutant p53-273H is associated with a high frequency of lung adenocarcinoma in transgenic mice. Oncogene 21: 7831–7838. doi: 10.1038/sj.onc.1205909
|
| [125] |
Susse S, Janz C, Janus F, et al. (2000) Role of heteroduplex joints in the functional interactions between human Rad51 and wild-type p53. Oncogene 19: 4500–4512. doi: 10.1038/sj.onc.1203809
|
| [126] |
Subramanian D, Griffith JD (2002) Interactions between p53, hMSH2-hMSH6 and HMG I(Y) on Holliday junctions and bulged bases. Nucleic Acids Res 30: 2427–2434. doi: 10.1093/nar/30.11.2427
|
| [127] |
Kolinjivadi AM, Sannino V, de Antoni A, et al. (2017) Moonlighting at replication forks-a new life for homologous recombination proteins BRCA1, BRCA2 and RAD51. FEBS Lett 591: 1083–1100. doi: 10.1002/1873-3468.12556
|
| [128] |
Kolinjivadi AM, Sannino V, De Antoni A, et al. (2017) Smarcal1-Mediated Fork Reversal Triggers Mre11-Dependent Degradation of Nascent DNA in the Absence of Brca2 and Stable Rad51 Nucleofilaments. Mol Cell 67: 867–881. doi: 10.1016/j.molcel.2017.07.001
|
| [129] |
Lemacon D, Jackson J, Quinet A, et al. (2017) MRE11 and EXO1 nucleases degrade reversed forks and elicit MUS81-dependent fork rescue in BRCA2-deficient cells. Nat Commun 8: 860. doi: 10.1038/s41467-017-01180-5
|
| [130] |
Hashimoto Y, Chaudhuri AR, Lopes M, et al. (2010) Rad51 protects nascent DNA from Mre11-dependent degradation and promotes continuous DNA synthesis. Nat Struct Mol Biol 17: 1305–1311. doi: 10.1038/nsmb.1927
|
| [131] |
Suzuki A, de la Pompa JL, Hakem R, et al. (1997) Brca2 is required for embryonic cellular proliferation in the mouse. Genes Dev 11: 1242–1252. doi: 10.1101/gad.11.10.1242
|
| [132] |
Wang X, Takenaka K, Takeda S (2010) PTIP promotes DNA double-strand break repair through homologous recombination. Genes Cells 15: 243–254. doi: 10.1111/j.1365-2443.2009.01379.x
|
| [133] |
Ying S, Hamdy FC, Helleday T (2012) Mre11-dependent degradation of stalled DNA replication forks is prevented by BRCA2 and PARP1. Cancer Res 72: 2814–2821. doi: 10.1158/0008-5472.CAN-11-3417
|
| [134] |
Mijic S, Zellweger R, Chappidi N, et al. (2017) Replication fork reversal triggers fork degradation in BRCA2-defective cells. Nat Commun 8: 859. doi: 10.1038/s41467-017-01164-5
|
| [135] |
Ray Chaudhuri A, Callen E, Ding X, et al. (2016) Replication fork stability confers chemoresistance in BRCA-deficient cells. Nature 535: 382–387. doi: 10.1038/nature18325
|
| [136] |
Ding X, Ray Chaudhuri A, Callen E, et al. (2016) Synthetic viability by BRCA2 and PARP1/ARTD1 deficiencies. Nat Commun 7: 12425. doi: 10.1038/ncomms12425
|
| [137] |
Bhat KP, Krishnamoorthy A, Dungrawala H, et al. (2018) RADX modulates RAD51 activity to control replication fork protection. Cell reports 24: 538–545. doi: 10.1016/j.celrep.2018.06.061
|
| [138] |
Schubert L, Ho T, Hoffmann S, et al. (2017) RADX interacts with single-stranded DNA to promote replication fork stability. EMBO Rep 18: 1991–2003. doi: 10.15252/embr.201744877
|
| [139] |
Dungrawala H, Bhat KP, Le Meur R, et al. (2017) RADX promotes genome stability and modulates chemosensitivity by regulating RAD51 at replication forks. Mol Cell 67: 374–386. doi: 10.1016/j.molcel.2017.06.023
|
| [140] |
Taglialatela A, Alvarez S, Leuzzi G, et al. (2017) Restoration of replication fork stability in BRCA1- and BRCA2-deficient cells by inactivation of SNF2-family fork remodelers. Mol Cell 68: 414–430. doi: 10.1016/j.molcel.2017.09.036
|
| [141] |
Vujanovic M, Krietsch J, Raso MC, et al. (2017) Replication fork slowing and reversal upon DNA damage require PCNA polyubiquitination and ZRANB3 DNA translocase activity. Mol Cell 67: 882–890. doi: 10.1016/j.molcel.2017.08.010
|
| [142] |
Ciccia A, Bredemeyer AL, Sowa ME, et al. (2009) The SIOD disorder protein SMARCAL1 is an RPA-interacting protein involved in replication fork restart. Genes Dev 23: 2415–2425. doi: 10.1101/gad.1832309
|
| [143] |
Bansbach CE, Betous R, Lovejoy CA, et al. (2009) The annealing helicase SMARCAL1 maintains genome integrity at stalled replication forks. Genes Dev 23: 2405–2414. doi: 10.1101/gad.1839909
|
| [144] |
Postow L, Woo EM, Chait BT, et al. (2009) Identification of SMARCAL1 as a component of the DNA damage response. J Biol Chem 284: 35951–35961. doi: 10.1074/jbc.M109.048330
|
| [145] |
Yusufzai T, Kong X, Yokomori K, et al. (2009) The annealing helicase HARP is recruited to DNA repair sites via an interaction with RPA. Genes Dev 23: 2400–2404. doi: 10.1101/gad.1831509
|
| [146] |
Yuan J, Ghosal G, Chen J (2009) The annealing helicase HARP protects stalled replication forks. Genes Dev 23: 2394–2399. doi: 10.1101/gad.1836409
|
| [147] |
Yusufzai T, Kadonaga JT (2008) HARP is an ATP-driven annealing helicase. Science 322: 748–750. doi: 10.1126/science.1161233
|
| [148] |
Betous R, Mason AC, Rambo RP, et al. (2012) SMARCAL1 catalyzes fork regression and Holliday junction migration to maintain genome stability during DNA replication. Genes Dev 26: 151–162. doi: 10.1101/gad.178459.111
|
| [149] | Achar YJ, Balogh D, Neculai D, et al. (2015) Human HLTF mediates postreplication repair by its HIRAN domain-dependent replication fork remodelling. Nucleic Acids Res 43: 10277–10291. |
| [150] |
Berti M, Vindigni A (2016) Replication stress: Getting back on track. Nat Struct Mol Biol 23: 103–109. doi: 10.1038/nsmb.3163
|
| [151] |
Blastyak A, Hajdu I, Unk I, et al. (2010) Role of double-stranded DNA translocase activity of human HLTF in replication of damaged DNA. Mol Cell Biol 30: 684–693. doi: 10.1128/MCB.00863-09
|
| [152] |
Kile AC, Chavez DA, Bacal J, et al. (2015) HLTF's ancient HIRAN domain binds 3' DNA ends to drive replication fork reversal. Mol Cell 58: 1090–1100. doi: 10.1016/j.molcel.2015.05.013
|
| [153] |
Unk I, Hajdu I, Blastyak A, et al. (2010) Role of yeast Rad5 and its human orthologs, HLTF and SHPRH in DNA damage tolerance. DNA Repair 9: 257–267. doi: 10.1016/j.dnarep.2009.12.013
|
| [154] |
Motegi A, Liaw HJ, Lee KY, et al. (2008) Polyubiquitination of proliferating cell nuclear antigen by HLTF and SHPRH prevents genomic instability from stalled replication forks. Proc Natl Acad Sci USA 105: 12411–12416. doi: 10.1073/pnas.0805685105
|
| [155] |
Krijger PH, Lee KY, Wit N, et al. (2011) HLTF and SHPRH are not essential for PCNA polyubiquitination, survival and somatic hypermutation: Existence of an alternative E3 ligase. DNA Repair 10: 438–444. doi: 10.1016/j.dnarep.2010.12.008
|
| [156] |
Ulrich HD (2009) Regulating post-translational modifications of the eukaryotic replication clamp PCNA. DNA Repair 8: 461–469. doi: 10.1016/j.dnarep.2009.01.006
|
| [157] | Lee KY, Myung K (2008) PCNA modifications for regulation of post-replication repair pathways. Mol Cells 26: 5–11. |
| [158] |
McCulloch SD, Kunkel TA (2008) The fidelity of DNA synthesis by eukaryotic replicative and translesion synthesis polymerases. Cell Res 18: 148–161. doi: 10.1038/cr.2008.4
|
| [159] |
Badu-Nkansah A, Mason AC, Eichman BF, et al. (2016) Identification of a substrate recognition domain in the replication stress response protein Zinc finger Ran-binding domain-containing Protein 3 (ZRANB3). J Biol Chem 291: 8251–8257. doi: 10.1074/jbc.M115.709733
|
| [160] |
Weston R, Peeters H, Ahel D (2012) ZRANB3 is a structure-specific ATP-dependent endonuclease involved in replication stress response. Genes Dev 26: 1558–1572. doi: 10.1101/gad.193516.112
|
| [161] |
Lieber MR, Ma Y, Pannicke U, et al. (2003) Mechanism and regulation of human non-homologous DNA end-joining. Nat Rev Mol Cell Biol 4: 712–720. doi: 10.1038/nrm1202
|
| [162] |
Sallmyr A, Tomkinson AE (2018) Repair of DNA double-strand breaks by mammalian alternative end-joining pathways. J Biol Chem 293: 10536–10546. doi: 10.1074/jbc.TM117.000375
|
| [163] |
Llorente B, Smith CE, Symington LS (2008) Break-induced replication: What is it and what is it for? Cell Cycle 7: 859–864. doi: 10.4161/cc.7.7.5613
|
| [164] | Kurnit KC, Coleman RL, Westin SN (2018) Using PARP inhibitors in the treatment of patients with ovarian cancer. Curr Treat Options Oncol 19: 1. |
| [165] |
Farmer H, McCabe N, Lord CJ, et al. (2005) Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature 434: 917–921. doi: 10.1038/nature03445
|
| [166] |
Drean A, Lord CJ, Ashworth A (2016) PARP inhibitor combination therapy. Crit Rev Oncol Hematol 108: 73–85. doi: 10.1016/j.critrevonc.2016.10.010
|
| [167] |
Zhu J, Zhou L, Wu G, et al. (2013) A novel small molecule RAD51 inactivator overcomes imatinib-resistance in chronic myeloid leukaemia. EMBO Mol Med 5: 353–365. doi: 10.1002/emmm.201201760
|
| [168] |
An Z, Yu JR, Park WY (2016) T0070907 inhibits repair of radiation-induced DNA damage by targeting RAD51. Toxicol In Vitro 37: 1–8. doi: 10.1016/j.tiv.2016.08.009
|
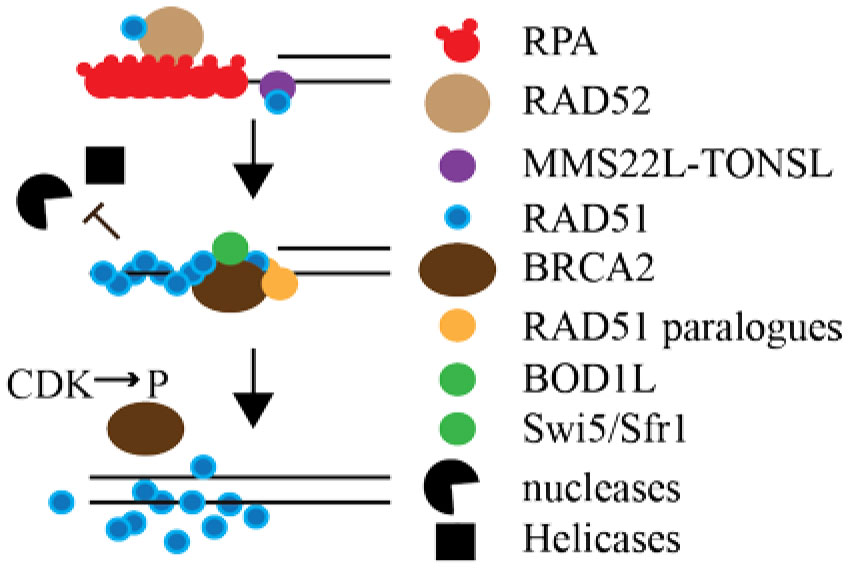
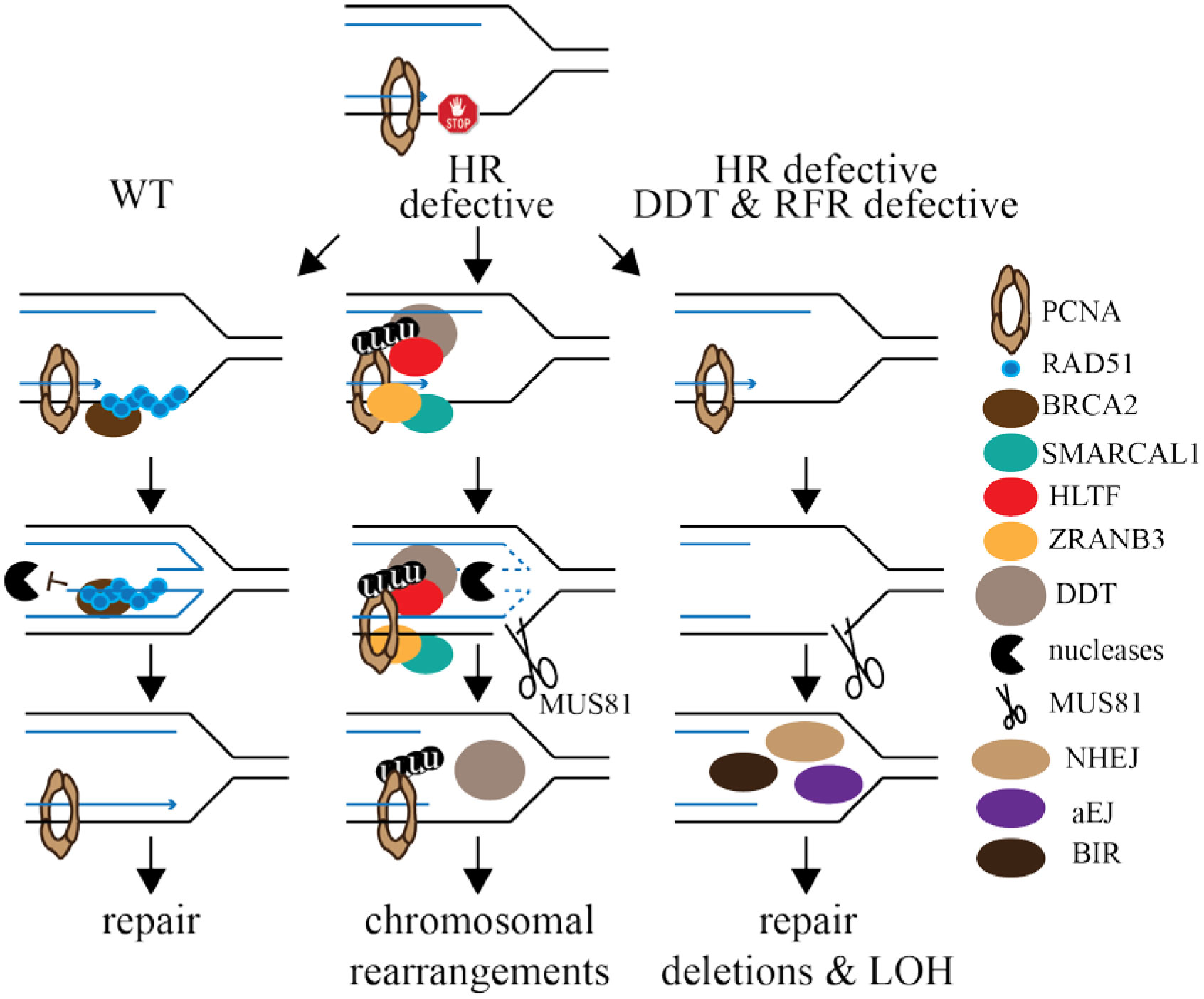
Figures(2)

Mi Young Son, Paul Hasty. Homologous recombination defects and how they affect replication fork maintenance[J]. AIMS Genetics, 2018, 5(4): 192-211. doi: 10.3934/genet.2018.4.192







 DownLoad:
DownLoad: