Citation: Mohsen Ahmadi Lashaki, Shahram Sedaghathoor, Sepideh Kalatehjari, Davood Hashemabadi. The physiological and growth response of Petunia hybrida, Tagetes erecta, and Calendula officinalis to plant and human steroids[J]. AIMS Agriculture and Food, 2018, 3(2): 85-96. doi: 10.3934/agrfood.2018.2.85

| [1] | Qasemi Gahsareh M, Kafi M (2007) Scientific and applied floriculture, Tehran: Golbol Press. |
| [2] |
Assar AWA, Echini KM (2011) Alleviation of drought stress of marigold (Tagetes erecta) plants by using arbuscular mycorrhizal fungi. Saudi J Biol Sci 18: 93–98. doi: 10.1016/j.sjbs.2010.06.007

|
| [3] | Omidbeigi R (2003) Production and processing of medicinal herbs, Mashhad: Astan-e Qods-e Razavi Press. |
| [4] |
Jones JL, Roddick JG (1988) Steroidal oestrogens and androgens in relation toreproductive development in higher plants. J Plant Physiol 133: 510–518. doi: 10.1016/S0176-1617(88)80047-6
|
| [5] |
Geuns JMC (1978) Steroid hormones and plant growth and development. Phytochemistry 17: 1–14. doi: 10.1016/S0031-9422(00)89671-5
|
| [6] | Ahmadi Mousavi E, Kalantari M, Torkzadeh M (2005) Effects of 24-epibrassinolide on lipid peroxidation, prolin, sugar and photosynthesis pigments content of rape (Brassica napus L.) under water stress. Iran J Biol 18: 295–306. |
| [7] | Janeczko A, Skocczowski A (2005) Mammalian sex hormones in plants. Folia Histochem Cytobiol 43: 71–79. |
| [8] | Zhang JS, Yang ZH, Tsao TH (1991) The occurrence of estrogens in relation to reproductive processes in flowering plants. Sex Plant Reprod 4: 193–196. |
| [9] |
Gendron JM, Wang ZY (2007) Multiple mechanisms modulate brassinosteroid signaling. Curr Opin Plant Biol 10:436–441. doi: 10.1016/j.pbi.2007.08.015
|
| [10] | Fahimi H (2008) Plant growth regulators , 2Eds., Tehran: Tehran University Press. |
| [11] | Arteca RN (2013) Plant growth substances: Principles and applications. Springer Sci Business Media. |
| [12] |
Bishop GJ, Koncz C (2002) Brassinosteroids and plant steroid hormone signaling. Plant Cell 14: S97–S110. doi: 10.1105/tpc.001461
|
| [13] |
Simons RG, Grinwich DL (1989) Immunoreactive detection of four mammalian steroids in plants. Can J Bot 67: 288–296. doi: 10.1139/b89-042
|
| [14] |
Milanesi L, Monje P, Boland R (2001) Presence of estrogens and estrogen receptor-likeproteins in Solanum glaucophyllum. Biochem Biophys Res Commun 289: 1175–1179. doi: 10.1006/bbrc.2001.6079
|
| [15] | Kopcewicz J (1969) Effect of estrone on the content of endogenous gibberellins in the dwarf pea. Naturwissenschaften 56: 334. |
| [16] |
Bhattacharya B, Gupta K (1981) Steroid hormone effects on growth and apical dominance of sunflower. Phytochemistry 20: 989–991. doi: 10.1016/0031-9422(81)83014-2
|
| [17] |
Sathiyabama M, Bernstein N, Anusuya S (2016) Chitosan elicitation for increased curcumin production and stimulation of defense response in turmeric (Curcuma longa L.). Ind Crops Prod 89: 87–94. doi: 10.1016/j.indcrop.2016.05.007
|
| [18] |
Gorelick J, Bernstein N (2014) Elicitation: An underutilized tool for the development of medicinal plants as a source for therapeutic secondary metabolites. Adv Agron 124: 201–230. doi: 10.1016/B978-0-12-800138-7.00005-X
|
| [19] |
Moll RH, Kamparth EJ (1977) Effect of population density up on agronomic traits associated with genetic increases in yield of (Zea mays L.). Agron J 69: 81–84. doi: 10.2134/agronj1977.00021962006900010021x
|
| [20] | Mazumdar BC, Majumder K (2003) Methods on physcochemical analysis of fruits. Calcutta: University College of Agriculture. 136–150. |
| [21] | Addy SK, Goodman RN (1972) Polyphenol oxidase and peroxidase in apple leaves inoculated with a virulent or an avirulent strain for Ervinia amylovora. Indian Phytopathol 25: 575–579. |
| [22] |
Dazy M, Jung V, Ferard J, et al. (2008) Ecological recovery of vegetation on a coke-factory soil: Role of plant antioxidant enzymes and possible implication in site restoration. Chemosphere 74: 57–63. doi: 10.1016/j.chemosphere.2008.09.014
|
| [23] | Das K, Samanta L, Chainy GBN (2000) A modified spectrophotometric assay of superoxide dismutase using nitrite formation by superoxide radicals. Indian J Biochem Biophys 37: 201–204. |
| [24] |
Lizaso JI, Batchelor WD, Westgate ME (2003) A leaf area model to simulate cultivar-specific expansion and senescence of maize leaves. Field Crops Res 80: 1–17. doi: 10.1016/S0378-4290(02)00151-X
|
| [25] |
Cristofori V, Rouphael Y, Gyves ME, et al. (2007) A simple model for estimating leaf area of hazelnut from linear measurements. Sci Hortic 113: 221–225. doi: 10.1016/j.scienta.2007.02.006
|
| [26] |
Anjum SA, Wang LC, Farooq M, et al. (2011) Brassinolide application improves the drought tolerancein maize through modulation of enzymaticantioxidants and leaf gas exchange. J Agron Crop Sci 197: 177–185. doi: 10.1111/j.1439-037X.2010.00459.x
|
| [27] | Prakash M, Suganthi S, Gokulakrishnan J, et al. (2008) Effect of homobrassinolide on growth, physiology and biochemical aspects of sesame. Karnataka J Agric Sci 20: 110–112. |
| [28] | Pasyar R, Saffari V (2014) The effect of 24-epibrassinolide on morphological traits and photosynthesis pigments of Gazania splendens L. Proceedings of the First National Conference of Flowers and Ornamental Plants of Iran, Karaj, Iran. |
| [29] | Shabani Soltanmoradi F (2016) A study on the effect of benzyladenine and epibrassinolide on the growth and development of Calendula officinalis L. (M.Sc. Thesis). Rasht Branch, Islamic Azad University, Rasht, Iran. |
| [30] |
Bakó E, Deli J, Tóth G (2002) HPLC study on the carotenoid composition of Calendula products. J Biochem Biophys Methods 53: 241–250. doi: 10.1016/S0165-022X(02)00112-4
|
| [31] |
Castenmiller JJM, West CE (1998) Bioavailability and bioconversion of carotenoids. Annu Rev Nutr 18: 19–38. doi: 10.1146/annurev.nutr.18.1.19
|
| [32] | Goodwin TW, Britton G, (1988) Distribution and analysis of carotenoids, In: Goodman TW, ed., Plant Pigments, London: Academic Press, 62–132. |
| [33] |
Khachik F, Steck A, Pfander H (1999) Isolation and structural elucidation of (13Z, 13'Z, 3R, 3'R, 6'R)-lutein from marigold flowers, kale, and human plasma. J Agric Food Chem 47: 455–461. doi: 10.1021/jf980823w
|
| [34] | Fariduddin Q, Yusuf M, Chalkoo S, et al. (2011) 28-homobrassinolide improves growth and photosynthesis in (Cucumis sativus L.) through an enhanced antioxidant system in the presence of chilling stress. Photosynthetica 49: 55–64. |
| [35] | Behnamnia M, Kalantariand KM, Rezanejad F (2009) Exogenous application of brassinosteroid alleviates drought-induced oxidative estress in (Lycopersicon esculentum L.). Bulg J Plant Physiol 35: 22–34. |
| [36] | Sardoei Kara Z, Saffari V, Tavassolian I (2016) Investigation of the response of morphological, physiological and biochemica attributes of marigold to seed priming and 24-epibrassinolide foliar application. Agric Crop Manag 18: 557–567. |
| [37] |
Hayat S, Maheshwari P, Wani AS, et al. (2012) Comparative effect of 28-homobrassinolide and salicylic acid in the amelioration of NaCl stress in Brassica juncea L. Plant Physiol Biochem 53: 61–68. doi: 10.1016/j.plaphy.2012.01.011
|
| [38] | Sadeghi F, Shekafandeh A (2014) Effect of 24-epibrassinolide on salinity-induced changes in loquat (Eriobotrya japonica Lindl). J Appl Bot Food Qual 87: 182–189. |
| [39] |
Swamy KN, Rao SSR (2006) Influence of brassinostroids on rooting and growth of geranium (Pelargonium sp.) stem cuttings. Asian J Plant Sci 5: 619–622. doi: 10.3923/ajps.2006.619.622
|
| [40] | Sarvajeet SG, Narendra T (2010) Reactive oxygen species and antioxidant machinery in a biotic stress tolerance in crop plants. Plant Physiol Biochem 3: 1–22. |
| [41] |
Yuan GF, Jia CG, Li Z, et al. (2010) Effect ofbrassinosteroids on drought resistance andabscisic acid concentration in tomato under waterstress. Sci Hortic 126: 103–108. doi: 10.1016/j.scienta.2010.06.014
|
| [42] |
Wu Y, Taylor KE, Biswas N, et al. (1999) Kinetic model for removal of phenol by horseradish peroxidase with PEG. J Environ Eng 125: 451–458. doi: 10.1061/(ASCE)0733-9372(1999)125:5(451)
|
| [43] |
Yamasaki H, Sakihama Y, Ikehara N (1997) Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiol 115: 1405–1412. doi: 10.1104/pp.115.4.1405
|
| [44] |
Li KR, Wang HH, Han G, et al. (2008) Effects of brassinolide on the survival, growth and drought resistance of Robinia pseudoacaciaseedlings under water-stress. New Forests 35: 255–266. doi: 10.1007/s11056-007-9075-2
|
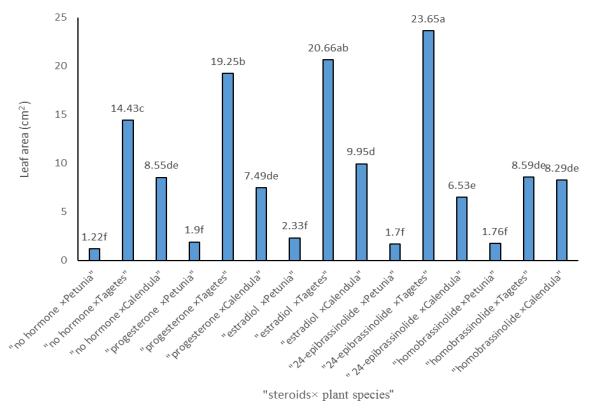
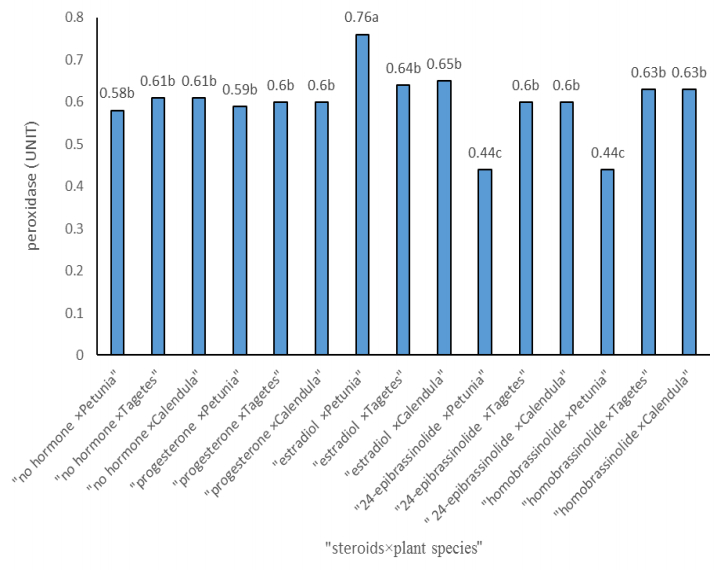
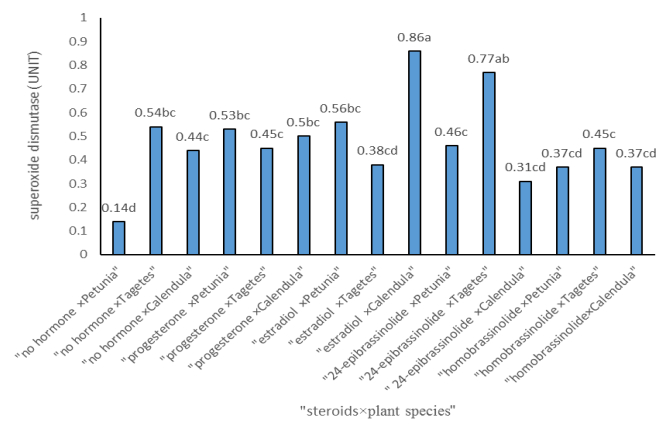
Figures(3) / Tables(3)

Mohsen Ahmadi Lashaki, Shahram Sedaghathoor, Sepideh Kalatehjari, Davood Hashemabadi. The physiological and growth response of Petunia hybrida, Tagetes erecta, and Calendula officinalis to plant and human steroids[J]. AIMS Agriculture and Food, 2018, 3(2): 85-96. doi: 10.3934/agrfood.2018.2.85







 DownLoad:
DownLoad: