Citation: Fatima Al-Hashimi, Salvador J. Diaz-Cano. Multi-target analysis of neoplasms for the evaluation of tumor progression: stochastic approach of biologic processes[J]. AIMS Molecular Science, 2018, 5(1): 14-62. doi: 10.3934/molsci.2018.1.14

| [1] | Diaz-Cano SJ (2008) General morphological and biological features of neoplasms: Integration of molecular findings. Histopathology 53: 1–19. |
| [2] |
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: The next generation. Cell 144: 646–674. doi: 10.1016/j.cell.2011.02.013

|
| [3] |
Hanahan D, Weinberg RA (2000) The hallmarks of cancer. Cell 100: 57–70. doi: 10.1016/S0092-8674(00)81683-9
|
| [4] |
Kinzler KW, Vogelstein B (1996) Lessons from hereditary colorectal cancer. Cell 87: 159–170. doi: 10.1016/S0092-8674(00)81333-1
|
| [5] |
Fidler IJ, Hart IR (1982) Biological diversity in metastatic neoplasms: Origins and implications. Science 217: 998–1003. doi: 10.1126/science.7112116
|
| [6] | Heppner GH, Miller FR (1998) The cellular basis of tumor progression. Int Rev Cytol 177: 1–56. |
| [7] |
Talmadge JE, Wolman SR, Fidler IJ (1982) Evidence for the clonal origin of spontaneous metastases. Science 217: 361–363. doi: 10.1126/science.6953592
|
| [8] |
Blanes A, Diaz-Cano SJ (2006) Complementary analysis of microsatellite tumor profile and mismatch repair defects in colorectal carcinomas. World J Gastroenterol 12: 5932–5940. doi: 10.3748/wjg.v12.i37.5932
|
| [9] |
Blanes A, Diaz-Cano SJ (2006) DNA and kinetic heterogeneity during the clonal evolution of adrenocortical proliferative lesions. Hum Pathol 37: 1295–1303. doi: 10.1016/j.humpath.2006.04.025
|
| [10] |
Blanes A, Sanchez-Carrillo JJ, Diaz-Cano SJ (2006) Topographic molecular profile of pheochromocytomas: Role of somatic down-regulation of mismatch repair. J Clin Endocrinol Metab 91: 1150–1158. doi: 10.1210/jc.2005-1645
|
| [11] | Pozo L, Sanchez-Carrillo JJ, Martinez A, et al. (2007) Differential kinetic features by tumour topography in cutaneous small-cell neuroendocrine (Merkel cell) carcinomas. J Eur Acad Dermatol Venereol 21: 1220–1228. |
| [12] |
Kinzler KW, Vogelstein B (1998) Landscaping the cancer terrain. Science 280: 1036–1037. doi: 10.1126/science.280.5366.1036
|
| [13] |
Parmigiani G, Boca S, Lin J, et al. (2009) Design and analysis issues in genome-wide somatic mutation studies of cancer. Genomics 93: 17–21. doi: 10.1016/j.ygeno.2008.07.005
|
| [14] |
Polyak K, Haviv I, Campbell IG (2009) Co-evolution of tumor cells and their microenvironment. Trends Genet 25: 30–38. doi: 10.1016/j.tig.2008.10.012
|
| [15] |
Nowell PC (1976) The clonal evolution of tumor cell populations. Science 194: 23–28. doi: 10.1126/science.959840
|
| [16] |
McGranahan N, Swanton C (2017) Clonal Heterogeneity and Tumor Evolution: Past, Present, and the Future. Cell 168: 613–628. doi: 10.1016/j.cell.2017.01.018
|
| [17] |
Shah NP, Nicoll JM, Nagar B, et al. (2002) Multiple BCR-ABL kinase domain mutations confer polyclonal resistance to the tyrosine kinase inhibitor imatinib (STI571) in chronic phase and blast crisis chronic myeloid leukemia. Cancer Cell 2: 117–125. doi: 10.1016/S1535-6108(02)00096-X
|
| [18] |
Corless CL, Heinrich MC (2008) Molecular pathobiology of gastrointestinal stromal sarcomas. Annu Rev Pathol 3: 557–586. doi: 10.1146/annurev.pathmechdis.3.121806.151538
|
| [19] |
Edwards SL, Brough R, Lord CJ, et al. (2008) Resistance to therapy caused by intragenic deletion in BRCA2. Nature 451: 1111–1115. doi: 10.1038/nature06548
|
| [20] |
Blagosklonny MV (2002) Oncogenic resistance to growth-limiting conditions. Nat Rev Cancer 2: 221–225. doi: 10.1038/nrc743
|
| [21] |
Laconi E, Doratiotto S, Vineis P (2008) The microenvironments of multistage carcinogenesis. Semin Cancer Biol 18: 322–329. doi: 10.1016/j.semcancer.2008.03.019
|
| [22] |
Laconi E, Sonnenschein C (2008) Cancer development at tissue level. Semin Cancer Biol 18: 303–304. doi: 10.1016/j.semcancer.2008.03.002
|
| [23] | Bissell MJ, Labarge MA (2005) Context, tissue plasticity, and cancer: Are tumor stem cells also regulated by the microenvironment? Cancer Cell 7: 17–23. |
| [24] |
Polyak K (2011) Heterogeneity in breast cancer. J Clin Invest 121: 3786–3788. doi: 10.1172/JCI60534
|
| [25] |
Merlo LM, Pepper JW, Reid BJ, et al. (2006) Cancer as an evolutionary and ecological process. Nat Rev Cancer 6: 924–935. doi: 10.1038/nrc2013
|
| [26] |
Axelrod R, Axelrod DE, Pienta KJ (2006) Evolution of cooperation among tumor cells. Proc Natl Acad Sci U S A 103: 13474–13479. doi: 10.1073/pnas.0606053103
|
| [27] |
Mallepell S, Krust A, Chambon P, et al. (2006) Paracrine signaling through the epithelial estrogen receptor alpha is required for proliferation and morphogenesis in the mammary gland. Proc Natl Acad Sci U S A 103: 2196–2201. doi: 10.1073/pnas.0510974103
|
| [28] |
Godar S, Ince TA, Bell GW, et al. (2008) Growth-inhibitory and tumor- suppressive functions of p53 depend on its repression of CD44 expression. Cell 134: 62–73. doi: 10.1016/j.cell.2008.06.006
|
| [29] |
McAllister SS, Gifford AM, Greiner AL, et al. (2008) Systemic endocrine instigation of indolent tumor growth requires osteopontin. Cell 133: 994–1005. doi: 10.1016/j.cell.2008.04.045
|
| [30] |
Nagy JD (2004) Competition and natural selection in a mathematical model of cancer. Bull Math Biol 66: 663–687. doi: 10.1016/j.bulm.2003.10.001
|
| [31] |
Yamamoto K, Hanada R, Kikuchi A, et al. (1998) Spontaneous regression of localized neuroblastoma detected by mass screening. J Clin Oncol 16: 1265–1269. doi: 10.1200/JCO.1998.16.4.1265
|
| [32] |
Axelrod R, Hamilton WD (1981) The evolution of cooperation. Science 211: 1390–1396. doi: 10.1126/science.7466396
|
| [33] |
Lyons JG, Lobo E, Martorana AM, et al. (2008) Clonal diversity in carcinomas: Its implications for tumour progression and the contribution made to it by epithelial-mesenchymal transitions. Clin Exp Metastasis 25: 665–677. doi: 10.1007/s10585-007-9134-2
|
| [34] | Marusyk A, DeGregori J (2008) Declining cellular fitness with age promotes cancer initiation by selecting for adaptive oncogenic mutations. Biochim Biophys Acta 1785: 1–11. |
| [35] |
Lowe SW, Cepero E, Evan G (2004) Intrinsic tumour suppression. Nature 432: 307–315. doi: 10.1038/nature03098
|
| [36] |
Zhang J, Grindley JC, Yin T, et al. (2006) PTEN maintains haematopoietic stem cells and acts in lineage choice and leukaemia prevention. Nature 441: 518–522. doi: 10.1038/nature04747
|
| [37] |
Michor F, Iwasa Y, Nowak MA (2004) Dynamics of cancer progression. Nat Rev Cancer 4: 197–205. doi: 10.1038/nrc1295
|
| [38] |
Michor F, Polyak K (2010) The origins and implications of intratumor heterogeneity. Cancer Prev Res 3: 1361–1364. doi: 10.1158/1940-6207.CAPR-10-0234
|
| [39] |
Marusyk A, Casas-Selves M, Henry CJ, et al. (2009) Irradiation alters selection for oncogenic mutations in hematopoietic progenitors. Cancer Res 69: 7262–7269. doi: 10.1158/0008-5472.CAN-09-0604
|
| [40] |
Laconi S, Pani P, Pillai S, et al. (2001) A growth-constrained environment drives tumor progression invivo. Proc Natl Acad Sci U S A 98: 7806–7811. doi: 10.1073/pnas.131210498
|
| [41] |
Devireddy LR, Gazin C, Zhu X, et al. (2005) A cell-surface receptor for lipocalin 24p3 selectively mediates apoptosis and iron uptake. Cell 123: 1293–1305. doi: 10.1016/j.cell.2005.10.027
|
| [42] | Guba M, Cernaianu G, Koehl G, et al. (2001) A primary tumor promotes dormancy of solitary tumor cells before inhibiting angiogenesis. Cancer Res 61: 5575–5579. |
| [43] |
Iwasa Y, Michor F (2011) Evolutionary dynamics of intratumor heterogeneity. PLoS One 6: e17866. doi: 10.1371/journal.pone.0017866
|
| [44] |
Diaz-Cano SJ (1998) Clonality studies in the analysis of adrenal medullary proliferations: Application principles and limitations. Endocr Pathol 9: 301–316. doi: 10.1007/BF02739690
|
| [45] |
Diaz-Cano SJ (2000) Designing a molecular analysis of clonality in tumours. J Pathol 191: 343–344. doi: 10.1002/1096-9896(200008)191:4<343::AID-PATH617>3.0.CO;2-Y
|
| [46] |
Diaz-Cano SJ, Blanes A, Wolfe HJ (2001) PCR techniques for clonality assays. Diagn Mol Pathol 10: 24–33. doi: 10.1097/00019606-200103000-00005
|
| [47] | Pozo-Garcia L, Diaz-Cano SJ (2003) Clonal origin and expansions in neoplasms: Biologic and technical aspects must be considered together. Am J Pathol 162: 354–355. |
| [48] |
Leedham SJ, Wright NA (2008) Human tumour clonality assessment-flawed but necessary. J Pathol 215: 351–354. doi: 10.1002/path.2379
|
| [49] |
Maley CC, Galipeau PC, Finley JC, et al. (2006) Genetic clonal diversity predicts progression to esophageal adenocarcinoma. Nat Genet 38: 468–473. doi: 10.1038/ng1768
|
| [50] |
Prasetyanti PR, Medema JP (2017) Intra-tumor heterogeneity from a cancer stem cell perspective. Mol Cancer 16: 41. doi: 10.1186/s12943-017-0600-4
|
| [51] |
Arif S, Blanes A, Diaz-Cano SJ (2002) Hashimoto's thyroiditis shares features with early papillary thyroid carcinoma. Histopathology 41: 357–362. doi: 10.1046/j.1365-2559.2002.01467.x
|
| [52] |
Blanes A, Rubio J, Martinez A, et al. (2002) Kinetic profiles by topographic compartments in muscle-invasive transitional cell carcinomas of the bladder: Role of TP53 and NF1 genes. Am J Clin Pathol 118: 93–100. doi: 10.1309/8QR9-2FRE-JPHF-VRC6
|
| [53] |
Diaz-Cano SJ, Blanes A, Rubio J, et al. (2000) Molecular evolution and intratumor heterogeneity by topographic compartments in muscle-invasive transitional cell carcinoma of the urinary bladder. Lab Invest 80: 279–289. doi: 10.1038/labinvest.3780033
|
| [54] |
Diaz-Cano SJ, Miguel MD, Blanes A, et al. (2000) Clonality as expression of distinctive cell kinetics patterns in nodular hyperplasias and adenomas of the adrenal cortex. Am J Pathol 156: 311–319. doi: 10.1016/S0002-9440(10)64732-3
|
| [55] |
Diaz-Cano SJ, Miguel MD, Blanes A, et al. (2000) Clonal patterns in phaeochromocytomas and MEN-2A adrenal medullary hyperplasias: Histological and kinetic correlates. J Pathol 192: 221–228. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH679>3.0.CO;2-B
|
| [56] | Jones PA (1986) DNA methylation and cancer. Cancer Res 46: 461–466. |
| [57] |
Jones PA, Baylin SB (2007) The epigenomics of cancer. Cell 128: 683–692. doi: 10.1016/j.cell.2007.01.029
|
| [58] |
Diaz-Cano SJ (2001) Are PCR artifacts in microdissected samples preventable? Hum Pathol 32: 1415–1416. doi: 10.1053/hupa.2001.29632
|
| [59] |
Diaz-Cano SJ, Miguel MD, Blanes A, et al. (2001) Contribution of the microvessel network to the clonal and kinetic profiles of adrenal cortical proliferative lesions. Hum Pathol 32: 1232–1239. doi: 10.1053/hupa.2001.28949
|
| [60] |
Zellmer VR, Zhang S (2014) Evolving concepts of tumor heterogeneity. Cell Biosci 4: 69. doi: 10.1186/2045-3701-4-69
|
| [61] |
Blanes A, Rubio J, Sanchez-Carrillo JJ, et al. (2009) Coexistent intraurothelial carcinoma and muscle-invasive urothelial carcinoma of the bladder: Clonality and somatic down-regulation of DNA mismatch repair. Hum Pathol 40: 988–997. doi: 10.1016/j.humpath.2008.12.009
|
| [62] |
Diaz-Cano SJ (2015) Pathological bases for a robust application of cancer molecular classification. Int J Mol Sci 16: 8655–8675. doi: 10.3390/ijms16048655
|
| [63] |
Diaz-Cano SJ, de Miguel M, Blanes A, et al. (2001) Germline RET 634 mutation positive MEN 2A-related C-cell hyperplasias have genetic features consistent with intraepithelial neoplasia. J Clin Endocrinol Metab 86: 3948–3957. doi: 10.1210/jcem.86.8.7739
|
| [64] |
Rubio J, Blanes A, Sanchez-Carrillo JJ, et al. (2007) Microsatellite abnormalities and somatic down-regulation of mismatch repair characterize nodular-trabecular muscle-invasive urothelial carcinoma of the bladder. Histopathology 51: 458–467. doi: 10.1111/j.1365-2559.2007.02795.x
|
| [65] |
Husain EA, Mein C, Pozo L, et al. (2011) Heterogeneous topographic profiles of kinetic and cell cycle regulator microsatellites in atypical (dysplastic) melanocytic nevi. Mod Pathol 24: 471–486. doi: 10.1038/modpathol.2010.143
|
| [66] |
Morrison SJ, Spradling AC (2008) Stem cells and niches: Mechanisms that promote stem cell maintenance throughout life. Cell 132: 598–611. doi: 10.1016/j.cell.2008.01.038
|
| [67] |
Reddig PJ, Juliano RL (2005) Clinging to life: Cell to matrix adhesion and cell survival. Cancer Metastasis Rev 24: 425–439. doi: 10.1007/s10555-005-5134-3
|
| [68] |
Tlsty TD, Coussens LM (2006) Tumor stroma and regulation of cancer development. Annu Rev Pathol 1: 119–150. doi: 10.1146/annurev.pathol.1.110304.100224
|
| [69] |
Baithun SI, Naase M, Blanes A, et al. (2001) Molecular and kinetic features of transitional cell carcinomas of the bladder: Biological and clinical implications. Virchows Arch 438: 289–297. doi: 10.1007/s004280000289
|
| [70] |
Durrett R, Foo J, Leder K, et al. (2011) Intratumor heterogeneity in evolutionary models of tumor progression. Genetics 188: 461–477. doi: 10.1534/genetics.110.125724
|
| [71] |
Laconi E (2007) The evolving concept of tumor microenvironments. Bioessays 29: 738–744. doi: 10.1002/bies.20606
|
| [72] |
Huang LE, Bindra RS, Glazer PM, et al. (2007) Hypoxia-induced genetic instability-a calculated mechanism underlying tumor progression. J Mol Med 85: 139–148. doi: 10.1007/s00109-006-0133-6
|
| [73] |
Hahn WC, Counter CM, Lundberg AS, et al. (1999) Creation of human tumour cells with defined genetic elements. Nature 400: 464–468. doi: 10.1038/22780
|
| [74] |
Rubin H (2003) Microenvironmental regulation of the initiated cell. Adv Cancer Res 90: 1–62. doi: 10.1016/S0065-230X(03)90001-7
|
| [75] |
Mudgil AV, Segal N, Andriani F, et al. (2003) Ultraviolet B irradiation induces expansion of intraepithelial tumor cells in a tissue model of early cancer progression. J Invest Dermatol 121: 191–197. doi: 10.1046/j.1523-1747.2003.12320.x
|
| [76] |
Schollnberger H, Manuguerra M, Bijwaard H, et al. (2006) Analysis of epidemiological cohort data on smoking effects and lung cancer with a multi-stage cancer model. Carcinogenesis 27: 1432–1444. doi: 10.1093/carcin/bgi345
|
| [77] | Farber E, Rubin H (1991) Cellular adaptation in the origin and development of cancer. Cancer Res 51: 2751–2761. |
| [78] |
Cristini V, Frieboes HB, Gatenby R, et al. (2005) Morphologic instability and cancer invasion. Clin Cancer Res 11: 6772–6779. doi: 10.1158/1078-0432.CCR-05-0852
|
| [79] |
Cayama E, Tsuda H, Sarma DS, et al. (1978) Initiation of chemical carcinogenesis requires cell proliferation. Nature 275: 60–62. doi: 10.1038/275060a0
|
| [80] |
Breivik J (2005) The evolutionary origin of genetic instability in cancer development. Semin Cancer Biol 15: 51–60. doi: 10.1016/j.semcancer.2004.09.008
|
| [81] |
West RB, Rubin BP, Miller MA, et al. (2006) A landscape effect in tenosynovial giant-cell tumor from activation of CSF1 expression by a translocation in a minority of tumor cells. Proc Natl Acad Sci U S A 103: 690–695. doi: 10.1073/pnas.0507321103
|
| [82] |
van Vugt MA, Bras A, Medema RH (2005) Restarting the cell cycle when the checkpoint comes to a halt. Cancer Res 65: 7037–7040. doi: 10.1158/0008-5472.CAN-05-1054
|
| [83] |
Jonason AS, Kunala S, Price GJ, et al. (1996) Frequent clones of p53-mutated keratinocytes in normal human skin. Proc Natl Acad Sci U S A 93: 14025–14029. doi: 10.1073/pnas.93.24.14025
|
| [84] |
Laconi E, Pani P, Farber E (2000) The resistance phenotype in the development and treatment of cancer. Lancet Oncol 1: 235–241. doi: 10.1016/S1470-2045(00)00154-6
|
| [85] |
Marongiu F, Doratiotto S, Montisci S, et al. (2008) Liver repopulation and carcinogenesis: Two sides of the same coin? Am J Pathol 172: 857–864. doi: 10.2353/ajpath.2008.070910
|
| [86] |
Campisi J (2007) Aging and cancer cell biology, 2007. Aging Cell 6: 261–263. doi: 10.1111/j.1474-9726.2007.00292.x
|
| [87] |
Sharpless NE, DePinho RA (2007) How stem cells age and why this makes us grow old. Nat Rev Mol Cell Biol 8: 703–713. doi: 10.1038/nrm2241
|
| [88] | McCullough KD, Coleman WB, Smith GJ, et al. (1994) Age-dependent regulation of the tumorigenic potential of neoplastically transformed rat liver epithelial cells by the liver microenvironment. Cancer Res 54: 3668–3671. |
| [89] |
Pasciu D, Montisci S, Greco M, et al. (2006) Aging is associated with increased clonogenic potential in rat liver in vivo. Aging Cell 5: 373–377. doi: 10.1111/j.1474-9726.2006.00230.x
|
| [90] |
Krtolica A, Parrinello S, Lockett S, et al. (2001) Senescent fibroblasts promote epithelial cell growth and tumorigenesis: A link between cancer and aging. Proc Natl Acad Sci U S A 98: 12072–12077. doi: 10.1073/pnas.211053698
|
| [91] |
Liu D, Hornsby PJ (2007) Senescent human fibroblasts increase the early growth of xenograft tumors via matrix metalloproteinase secretion. Cancer Res 67: 3117–3126. doi: 10.1158/0008-5472.CAN-06-3452
|
| [92] | Trost TM, Lausch EU, Fees SA, et al. (2005) Premature senescence is a primary fail-safe mechanism of ERBB2-driven tumorigenesis in breast carcinoma cells. Cancer Res 65: 840–849. |
| [93] |
Philip M, Rowley DA, Schreiber H (2004) Inflammation as a tumor promoter in cancer induction. Semin Cancer Biol 14: 433–439. doi: 10.1016/j.semcancer.2004.06.006
|
| [94] |
Hussain SP, Harris CC (2007) Inflammation and cancer: An ancient link with novel potentials. Int J Cancer 121: 2373–2380. doi: 10.1002/ijc.23173
|
| [95] |
Prehn RT (1997) Regeneration versus neoplastic growth. Carcinogenesis 18: 1439–1444. doi: 10.1093/carcin/18.8.1439
|
| [96] |
Correa P, Houghton J (2007) Carcinogenesis of Helicobacter pylori. Gastroenterology 133: 659–672. doi: 10.1053/j.gastro.2007.06.026
|
| [97] |
De Marzo AM, Marchi VL, Epstein JI, et al. (1999) Proliferative inflammatory atrophy of the prostate: Implications for prostatic carcinogenesis. Am J Pathol 155: 1985–1992. doi: 10.1016/S0002-9440(10)65517-4
|
| [98] |
De Marzo AM, Nakai Y, Nelson WG (2007) Inflammation, atrophy, and prostate carcinogenesis. Urol Oncol 25: 398–400. doi: 10.1016/j.urolonc.2007.05.007
|
| [99] |
Fox JG, Wang TC (2007) Inflammation, atrophy, and gastric cancer. J Clin Invest 117: 60–69. doi: 10.1172/JCI30111
|
| [100] |
Harpaz N (2007) Neoplastic precursor lesions related to the development of cancer in inflammatory bowel disease. Gastroenterol Clin North Am 36: 901–926, vii–viii. doi: 10.1016/j.gtc.2007.08.003
|
| [101] |
Rass U, Ahel I, West SC (2007) Defective DNA repair and neurodegenerative disease. Cell 130: 991–1004. doi: 10.1016/j.cell.2007.08.043
|
| [102] |
De Boer J, Andressoo JO, De Wit J, et al. (2002) Premature aging in mice deficient in DNA repair and transcription. Science 296: 1276–1279. doi: 10.1126/science.1070174
|
| [103] |
Simpson AJ (1997) The natural somatic mutation frequency and human carcinogenesis. Adv Cancer Res 71: 209–240. doi: 10.1016/S0065-230X(08)60100-1
|
| [104] |
Farber E, Cameron R (1980) The sequential analysis of cancer development. Adv Cancer Res 31: 125–226. doi: 10.1016/S0065-230X(08)60658-2
|
| [105] |
Bindra RS, Glazer PM (2005) Genetic instability and the tumor microenvironment: Owards the concept of microenvironment-induced mutagenesis. Mutat Res 569: 75–85. doi: 10.1016/j.mrfmmm.2004.03.013
|
| [106] |
Sneddon JB, Werb Z (2007) Location, location, location: The cancer stem cell niche. Cell Stem Cell 1: 607–611. doi: 10.1016/j.stem.2007.11.009
|
| [107] |
Calabrese C, Poppleton H, Kocak M, et al. (2007) A perivascular niche for brain tumor stem cells. Cancer Cell 11: 69–82. doi: 10.1016/j.ccr.2006.11.020
|
| [108] |
Veeravagu A, Bababeygy SR, Kalani MY, et al. (2008) The cancer stem cell-vascular niche complex in brain tumor formation. Stem Cells Dev 17: 859–867. doi: 10.1089/scd.2008.0047
|
| [109] |
Gilbertson RJ, Gutmann DH (2007) Tumorigenesis in the brain: Location, location, location. Cancer Res 67: 5579–5582. doi: 10.1158/0008-5472.CAN-07-0760
|
| [110] |
Gilbertson RJ, Rich JN (2007) Making a tumour's bed: Glioblastoma stem cells and the vascular niche. Nat Rev Cancer 7: 733–736. doi: 10.1038/nrc2246
|
| [111] |
Croker AK, Goodale D, Chu J, et al. (2009) High aldehyde dehydrogenase and expression of cancer stem cell markers selects for breast cancer cells with enhanced malignant and metastatic ability. J Cell Mol Med 13: 2236–2252. doi: 10.1111/j.1582-4934.2008.00455.x
|
| [112] |
Croker AK, Allan AL (2008) Cancer stem cells: Implications for the progression and treatment of metastatic disease. J Cell Mol Med 12: 374–390. doi: 10.1111/j.1582-4934.2007.00211.x
|
| [113] |
Campbell I, Qiu W, Haviv I (2011) Genetic changes in tumour microenvironments. J Pathol 223: 450–458. doi: 10.1002/path.2842
|
| [114] |
Korkaya H, Liu S, Wicha MS (2011) Breast cancer stem cells, cytokine networks, and the tumor microenvironment. J Clin Invest 121: 3804–3809. doi: 10.1172/JCI57099
|
| [115] |
Weinberg RA (2008) Coevolution in the tumor microenvironment. Nat Genet 40: 494–495. doi: 10.1038/ng0508-494
|
| [116] |
Whiteside TL (2008) The tumor microenvironment and its role in promoting tumor growth. Oncogene 27: 5904–5912. doi: 10.1038/onc.2008.271
|
| [117] |
Ben-Baruch A (2003) Host microenvironment in breast cancer development: Inflammatory cells, cytokines and chemokines in breast cancer progression: Reciprocal tumor-microenvironment interactions. Breast Cancer Res 5: 31–36. doi: 10.1186/bcr690
|
| [118] |
Zipin-Roitman A, Meshel T, Sagi-Assif O, et al. (2007) CXCL10 promotes invasion-related properties in human colorectal carcinoma cells. Cancer Res 67: 3396–3405. doi: 10.1158/0008-5472.CAN-06-3087
|
| [119] |
Malumbres M, Barbacid M (2001) To cycle or not to cycle: A critical decision in cancer. Nat Rev Cancer 1: 222–231. doi: 10.1038/35106065
|
| [120] |
Hersey P, Zhang XD (2001) How melanoma cells evade trail-induced apoptosis. Nat Rev Cancer 1: 142–150. doi: 10.1038/35101078
|
| [121] |
Anderson AR, Hassanein M, Branch KM, et al. (2009) Microenvironmental independence associated with tumor progression. Cancer Res 69: 8797–8806. doi: 10.1158/0008-5472.CAN-09-0437
|
| [122] |
Anderson AR, Rejniak KA, Gerlee P, et al. (2009) Microenvironment driven invasion: A multiscale multimodel investigation. J Math Biol 58: 579–624. doi: 10.1007/s00285-008-0210-2
|
| [123] |
Anderson AR, Weaver AM, Cummings PT, et al. (2006) Tumor morphology and phenotypic evolution driven by selective pressure from the microenvironment. Cell 127: 905–915. doi: 10.1016/j.cell.2006.09.042
|
| [124] |
Coussens LM, Werb Z (2002) Inflammation and cancer. Nature 420: 860–867. doi: 10.1038/nature01322
|
| [125] |
Ellis LM, Hicklin DJ (2008) VEGF-targeted therapy: Mechanisms of anti-tumour activity. Nat Rev Cancer 8: 579–591. doi: 10.1038/nrc2403
|
| [126] |
Yang L, Huang J, Ren X, et al. (2008) Abrogation of TGF beta signaling in mammary carcinomas recruits Gr-1+CD11b+ myeloid cells that promote metastasis. Cancer Cell 13: 23–35. doi: 10.1016/j.ccr.2007.12.004
|
| [127] |
Gabrilovich D (2004) Mechanisms and functional significance of tumour-induced dendritic-cell defects. Nat Rev Immunol 4: 941–952. doi: 10.1038/nri1498
|
| [128] |
Marx J (2008) All in the stroma: Cancer's Cosa Nostra. Science 320: 38–41. doi: 10.1126/science.320.5872.38
|
| [129] |
Marx J (2008) Cancer's bulwark against immune attack: MDS cells. Science 319: 154–156. doi: 10.1126/science.319.5860.154
|
| [130] |
Nonaka K, Saio M, Suwa T, et al. (2008) Skewing the Th cell phenotype toward Th1 alters the maturation of tumor-infiltrating mononuclear phagocytes. J Leukoc Biol 84: 679–688. doi: 10.1189/jlb.1107729
|
| [131] |
Umemura N, Saio M, Suwa T, et al. (2008) Tumor-infiltrating myeloid-derived suppressor cells are pleiotropic-inflamed monocytes/macrophages that bear M1- and M2-type characteristics. J Leukoc Biol 83: 1136–1144. doi: 10.1189/jlb.0907611
|
| [132] |
Mantovani A, Sica A, Allavena P, et al. (2009) Tumor-associated macrophages and the related myeloid-derived suppressor cells as a paradigm of the diversity of macrophage activation. Hum Immunol 70: 325–330. doi: 10.1016/j.humimm.2009.02.008
|
| [133] |
Sica A, Larghi P, Mancino A, et al. (2008) Macrophage polarization in tumour progression. Semin Cancer Biol 18: 349–355. doi: 10.1016/j.semcancer.2008.03.004
|
| [134] |
Mantovani A, Sica A (2010) Macrophages, innate immunity and cancer: Balance, tolerance, and diversity. Curr Opin Immunol 22: 231–237. doi: 10.1016/j.coi.2010.01.009
|
| [135] |
Sinha P, Clements VK, Miller S, et al. (2005) Tumor immunity: A balancing act between T cell activation, macrophage activation and tumor-induced immune suppression. Cancer Immunol Immunother 54: 1137–1142. doi: 10.1007/s00262-005-0703-4
|
| [136] |
Sinha P, Clements VK, Ostrand-Rosenberg S (2005) Reduction of myeloid-derived suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J Immunol 174: 636–645. doi: 10.4049/jimmunol.174.2.636
|
| [137] |
Sinha P, Clements VK, Ostrand-Rosenberg S (2005) Interleukin-13-regulated M2 macrophages in combination with myeloid suppressor cells block immune surveillance against metastasis. Cancer Res 65: 11743–11751. doi: 10.1158/0008-5472.CAN-05-0045
|
| [138] |
Lewis CE, Pollard JW (2006) Distinct role of macrophages in different tumor microenvironments. Cancer Res 66: 605–612. doi: 10.1158/0008-5472.CAN-05-4005
|
| [139] |
Yang L, DeBusk LM, Fukuda K, et al. (2004) Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 6: 409–421. doi: 10.1016/j.ccr.2004.08.031
|
| [140] |
Sinha P, Clements VK, Bunt SK, et al. (2007) Cross-talk between myeloid-derived suppressor cells and macrophages subverts tumor immunity toward a type 2 response. J Immunol 179: 977–983. doi: 10.4049/jimmunol.179.2.977
|
| [141] |
Condeelis J, Pollard JW (2006) Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 124: 263–266. doi: 10.1016/j.cell.2006.01.007
|
| [142] |
Larsen M, Tazzyman S, Lund EL, et al. (2008) Hypoxia-induced secretion of macrophage migration-inhibitory factor from MCF-7 breast cancer cells is regulated in a hypoxia-inducible factor-independent manner. Cancer Lett 265: 239–249. doi: 10.1016/j.canlet.2008.02.012
|
| [143] |
Bosco MC, Puppo M, Blengio F, et al. (2008) Monocytes and dendritic cells in a hypoxic environment: Spotlights on chemotaxis and migration. Immunobiology 213: 733–749. doi: 10.1016/j.imbio.2008.07.031
|
| [144] |
Youn JI, Nagaraj S, Collazo M, et al. (2008) Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J Immunol 181: 5791–5802. doi: 10.4049/jimmunol.181.8.5791
|
| [145] |
Movahedi K, Guilliams M, Van den Bossche J, et al. (2008) Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell-suppressive activity. Blood 111: 4233–4244. doi: 10.1182/blood-2007-07-099226
|
| [146] |
Kusmartsev S, Gabrilovich DI (2003) Inhibition of myeloid cell differentiation in cancer: The role of reactive oxygen species. J Leukoc Biol 74: 186–196. doi: 10.1189/jlb.0103010
|
| [147] |
Sinha P, Clements VK, Fulton AM, et al. (2007) Prostaglandin E2 promotes tumor progression by inducing myeloid-derived suppressor cells. Cancer Res 67: 4507–4513. doi: 10.1158/0008-5472.CAN-06-4174
|
| [148] | Bronte V, Serafini P, Mazzoni A, et al. (2003) L-arginine metabolism in myeloid cells controls T-lymphocyte functions. Trends Immunol 24: 302–306. |
| [149] |
Almand B, Clark JI, Nikitina E, et al. (2001) Increased production of immature myeloid cells in cancer patients: A mechanism of immunosuppression in cancer. J Immunol 166: 678–689. doi: 10.4049/jimmunol.166.1.678
|
| [150] |
Baniyash M (2004) TCR zeta-chain downregulation: Curtailing an excessive inflammatory immune response. Nat Rev Immunol 4: 675–687. doi: 10.1038/nri1434
|
| [151] |
Bunt SK, Sinha P, Clements VK, et al. (2006) Inflammation induces myeloid-derived suppressor cells that facilitate tumor progression. J Immunol 176: 284–290. doi: 10.4049/jimmunol.176.1.284
|
| [152] |
Bunt SK, Yang L, Sinha P, et al. (2007) Reduced inflammation in the tumor microenvironment delays the accumulation of myeloid-derived suppressor cells and limits tumor progression. Cancer Res 67: 10019–10026. doi: 10.1158/0008-5472.CAN-07-2354
|
| [153] |
Bielas JH, Loeb LA (2005) Mutator phenotype in cancer: Timing and perspectives. Environ Mol Mutagen 45: 206–213. doi: 10.1002/em.20111
|
| [154] |
Bardelli A, Cahill DP, Lederer G, et al. (2001) Carcinogen-specific induction of genetic instability. Proc Natl Acad Sci U S A 98: 5770–5775. doi: 10.1073/pnas.081082898
|
| [155] |
Jimenez JJ, Blanes A, Diaz-Cano SJ (2003) Microsatellite instability in colon cancer. N Engl J Med 349: 1774–1776. doi: 10.1056/NEJM200310303491818
|
| [156] |
Diaz-Cano SJ (2006) Molecular mechanisms in melanoma. N Engl J Med 355: 1395. doi: 10.1056/NEJMc062093
|
| [157] |
Pozo L, Husein E, Blanes A, et al. (2008) The correlation of regression with the grade of dysplasia (atypia) in melanocytic naevi. Histopathology 52: 387–389. doi: 10.1111/j.1365-2559.2007.02880.x
|
| [158] |
Husemann Y, Geigl JB, Schubert F, et al. (2008) Systemic spread is an early step in breast cancer. Cancer Cell 13: 58–68. doi: 10.1016/j.ccr.2007.12.003
|
| [159] |
Arif S, Patel J, Blanes A, et al. (2007) Cytoarchitectural and kinetic features in the histological evaluation of follicular thyroid neoplasms. Histopathology 50: 750–763. doi: 10.1111/j.1365-2559.2007.02680.x
|
| [160] |
Boone CW, Kelloff GJ, Freedman LS (1993) Intraepithelial and postinvasive neoplasia as a stochastic continuum of clonal evolution, and its relationship to mechanisms of chemopreventive drug action. J Cell Biochem Suppl 53: 14–25. doi: 10.1002/jcb.240531104
|
| [161] |
Namba R, Maglione JE, Davis RR, et al. (2006) Heterogeneity of mammary lesions represent molecular differences. BMC Cancer 6: 275. doi: 10.1186/1471-2407-6-275
|
| [162] |
Steeg PS (2006) Tumor metastasis: Mechanistic insights and clinical challenges. Nat Med 12: 895–904. doi: 10.1038/nm1469
|
| [163] |
Steeg PS (2007) Cancer: Icromanagement of metastasis. Nature 449: 671–673. doi: 10.1038/449671a
|
| [164] |
Talmadge JE (2007) Clonal selection of metastasis within the life history of a tumor. Cancer Res 67: 11471–11475. doi: 10.1158/0008-5472.CAN-07-2496
|
| [165] |
Christofori G (2006) New signals from the invasive front. Nature 441: 444–450. doi: 10.1038/nature04872
|
| [166] |
Gupta GP, Massague J (2006) Cancer metastasis: Building a framework. Cell 127: 679–695. doi: 10.1016/j.cell.2006.11.001
|
| [167] | Nguyen DX (2011) Tracing the origins of metastasis. J Pathol 223: 195–204. |
| [168] | Nguyen DX, Massague J (2007) Genetic determinants of cancer metastasis. Nat Rev Genet 8: 341–352. |
| [169] |
Li F, Tiede B, Massague J, et al. (2007) Beyond tumorigenesis: Cancer stem cells in metastasis. Cell Res 17: 3–14. doi: 10.1038/sj.cr.7310118
|
| [170] |
Wicha MS (2006) Cancer stem cells and metastasis: Lethal seeds. Clin Cancer Res 12: 5606–5607. doi: 10.1158/1078-0432.CCR-06-1537
|
| [171] | Kaplan RN, Psaila B, Lyden D (2006) Bone marrow cells in the 'pre-metastatic niche': Within bone and beyond. Cancer Metastasis Rev 25: 521–529. |
| [172] |
Kaplan RN, Rafii S, Lyden D (2006) Preparing the "soil": The premetastatic niche. Cancer Res 66: 11089–11093. doi: 10.1158/0008-5472.CAN-06-2407
|
| [173] |
Kaplan RN, Riba RD, Zacharoulis S, et al. (2005) VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 438: 820–827. doi: 10.1038/nature04186
|
| [174] |
Bissig H, Richter J, Desper R, et al. (1999) Evaluation of the clonal relationship between primary and metastatic renal cell carcinoma by comparative genomic hybridization. Am J Pathol 155: 267–274. doi: 10.1016/S0002-9440(10)65120-6
|
| [175] |
Cheng L, Bostwick DG, Li G, et al. (1999) Allelic imbalance in the clonal evolution of prostate carcinoma. Cancer 85: 2017–2022. doi: 10.1002/(SICI)1097-0142(19990501)85:9%3C2017::AID-CNCR20%3E3.0.CO;2-V
|
| [176] | Kuukasjarvi T, Karhu R, Tanner M, et al. (1997) Genetic heterogeneity and clonal evolution underlying development of asynchronous metastasis in human breast cancer. Cancer Res 57: 1597–1604. |
| [177] | Ramaswamy S, Ross KN, Lander ES, et al. (2003) A molecular signature of metastasis in primary solid tumors. Nat Genet 33: 49–54. |
| [178] |
Weigelt B, Glas AM, Wessels LF, et al. (2003) Gene expression profiles of primary breast tumors maintained in distant metastases. Proc Natl Acad Sci U S A 100: 15901–15905. doi: 10.1073/pnas.2634067100
|
| [179] |
Weigelt B, Hu Z, He X, et al. (2005) Molecular portraits and 70-gene prognosis signature are preserved throughout the metastatic process of breast cancer. Cancer Res 65: 9155–9158. doi: 10.1158/0008-5472.CAN-05-2553
|
| [180] |
Liu W, Laitinen S, Khan S, et al. (2009) Copy number analysis indicates monoclonal origin of lethal metastatic prostate cancer. Nat Med 15: 559–565. doi: 10.1038/nm.1944
|
| [181] | Ruijter ET, van de Kaa CA, Schalken JA, et al. (1996) Histological grade heterogeneity in multifocal prostate cancer. Biological and clinical implications. J Pathol 180: 295–299. |
| [182] |
Alvarado C, Beitel LK, Sircar K, et al. (2005) Somatic mosaicism and cancer: A micro-genetic examination into the role of the androgen receptor gene in prostate cancer. Cancer Res 65: 8514–8518. doi: 10.1158/0008-5472.CAN-05-0399
|
| [183] |
Shah SP, Morin RD, Khattra J, et al. (2009) Mutational evolution in a lobular breast tumour profiled at single nucleotide resolution. Nature 461: 809–813. doi: 10.1038/nature08489
|
| [184] |
Holmgren L, O'Reilly MS, Folkman J (1995) Dormancy of micrometastases: Balanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat Med 1: 149–153. doi: 10.1038/nm0295-149
|
| [185] |
Braun S, Pantel K, Muller P, et al. (2000) Cytokeratin-positive cells in the bone marrow and survival of patients with stage I, II, or III breast cancer. N Engl J Med 342: 525–533. doi: 10.1056/NEJM200002243420801
|
| [186] |
Diaz-Cano SJ (2000) Bone marrow metastases in breast cancer. N Engl J Med 343: 577. doi: 10.1056/NEJM200008243430811
|
| [187] |
Passlick B, Izbicki JR, Kubuschok B, et al. (1994) Immunohistochemical assessment of individual tumor cells in lymph nodes of patients with non-small-cell lung cancer. J Clin Oncol 12: 1827–1832. doi: 10.1200/JCO.1994.12.9.1827
|
| [188] |
Schlimok G, Funke I, Holzmann B, et al. (1987) Micrometastatic cancer cells in bone marrow: In vitro detection with anti-cytokeratin and in vivo labeling with anti-17-1A monoclonal antibodies. Proc Natl Acad Sci U S A 84: 8672–8676. doi: 10.1073/pnas.84.23.8672
|
| [189] |
Smith BL (2000) Approaches to breast-cancer staging. N Engl J Med 342: 580–581. doi: 10.1056/NEJM200002243420809
|
| [190] |
Klein CA, Schmidt-Kittler O, Schardt JA, et al. (1999) Comparative genomic hybridization, loss of heterozygosity, and DNA sequence analysis of single cells. Proc Natl Acad Sci U S A 96: 4494–4499. doi: 10.1073/pnas.96.8.4494
|
| [191] |
Klein CA, Blankenstein TJ, Schmidt-Kittler O, et al. (2002) Genetic heterogeneity of single disseminated tumour cells in minimal residual cancer. Lancet 360: 683–689. doi: 10.1016/S0140-6736(02)09838-0
|
| [192] |
Schmidt-Kittler O, Ragg T, Daskalakis A, et al. (2003) From latent disseminated cells to overt metastasis: Genetic analysis of systemic breast cancer progression. Proc Natl Acad Sci U S A 100: 7737–7742. doi: 10.1073/pnas.1331931100
|
| [193] |
Bedenne L, Michel P, Bouche O, et al. (2007) Chemoradiation followed by surgery compared with chemoradiation alone in squamous cancer of the esophagus: FFCD 9102. J Clin Oncol 25: 1160–1168. doi: 10.1200/JCO.2005.04.7118
|
| [194] |
Stoecklein NH, Hosch SB, Bezler M, et al. (2008) Direct genetic analysis of single disseminated cancer cells for prediction of outcome and therapy selection in esophageal cancer. Cancer Cell 13: 441–453. doi: 10.1016/j.ccr.2008.04.005
|
| [195] |
Podsypanina K, Du YC, Jechlinger M, et al. (2008) Seeding and propagation of untransformed mouse mammary cells in the lung. Science 321: 1841–1844. doi: 10.1126/science.1161621
|
| [196] |
Gadi VK, Nelson JL (2007) Fetal microchimerism in women with breast cancer. Cancer Res 67: 9035–9038. doi: 10.1158/0008-5472.CAN-06-4209
|
| [197] |
Solakoglu O, Maierhofer C, Lahr G, et al. (2002) Heterogeneous proliferative potential of occult metastatic cells in bone marrow of patients with solid epithelial tumors. Proc Natl Acad Sci U S A 99: 2246–2251. doi: 10.1073/pnas.042372199
|
| [198] |
Gangnus R, Langer S, Breit E, et al. (2004) Genomic profiling of viable and proliferative micrometastatic cells from early-stage breast cancer patients. Clin Cancer Res 10: 3457–3464. doi: 10.1158/1078-0432.CCR-03-0818
|
| [199] |
Abbruzzese JL, Abbruzzese MC, Hess KR, et al. (1994) Unknown primary carcinoma: Natural history and prognostic factors in 657 consecutive patients. J Clin Oncol 12: 1272–1280. doi: 10.1200/JCO.1994.12.6.1272
|
| [200] |
Klein CA, Holzel D (2006) Systemic cancer progression and tumor dormancy: Mathematical models meet single cell genomics. Cell Cycle 5: 1788–1798. doi: 10.4161/cc.5.16.3097
|
| [201] |
Beerenwinkel N, Antal T, Dingli D, et al. (2007) Genetic progression and the waiting time to cancer. PLoS Comput Biol 3: 225. doi: 10.1371/journal.pcbi.0030225
|
| [202] |
Sieber OM, Tomlinson SR, Tomlinson IP (2005) Tissue, cell and stage specificity of (epi)mutations in cancers. Nat Rev Cancer 5: 649–655. doi: 10.1038/nrc1674
|
| [203] |
Fialkow PJ (1979) Clonal origin of human tumors. Annu Rev Med 30: 135–143. doi: 10.1146/annurev.me.30.020179.001031
|
| [204] | Weinberg RA (2007) The biology of cancer, New York: Garland Science, 796. |
| [205] |
Novelli MR, Williamson JA, Tomlinson IP, et al. (1996) Polyclonal origin of colonic adenomas in an XO/XY patient with FAP. Science 272: 1187–1190. doi: 10.1126/science.272.5265.1187
|
| [206] |
Leroi AM, Koufopanou V, Burt A (2003) Cancer selection. Nat Rev Cancer 3: 226–231. doi: 10.1038/nrc1016
|
| [207] |
Potten CS (1998) Stem cells in gastrointestinal epithelium: Numbers, characteristics and death. Philos Trans R Soc Lond 353: 821–830. doi: 10.1098/rstb.1998.0246
|
| [208] |
Friedberg EC (2003) DNA damage and repair. Nature 421: 436–440. doi: 10.1038/nature01408
|
| [209] |
Tomlinson I, Bodmer W (1999) Selection, the mutation rate and cancer: Ensuring that the tail does not wag the dog. Nat Med 5: 11–12. doi: 10.1038/4687
|
| [210] |
Wang TL, Rago C, Silliman N, et al. (2002) Prevalence of somatic alterations in the colorectal cancer cell genome. Proc Natl Acad Sci U S A 99: 3076–3080. doi: 10.1073/pnas.261714699
|
| [211] |
Bielas JH, Loeb KR, Rubin BP, et al. (2006) Human cancers express a mutator phenotype. Proc Natl Acad Sci U S A 103: 18238–18242. doi: 10.1073/pnas.0607057103
|
| [212] |
Loeb LA (2016) Human cancers express a mutator phenotype: Hypothesis, origin, and consequences. Cancer Res 76: 2057–2059. doi: 10.1158/0008-5472.CAN-16-0794
|
| [213] |
Diaz-Cano SJ (2007) Kinetic topographical heterogeneity in follicular thyroid neoplasms and growth patterns. Histopathology 51: 416–418. doi: 10.1111/j.1365-2559.2007.02778.x
|
| [214] |
Gonzalez-Garcia I, Sole RV, Costa J (2002) Metapopulation dynamics and spatial heterogeneity in cancer. Proc Natl Acad Sci U S A 99: 13085–13089. doi: 10.1073/pnas.202139299
|
| [215] |
Gatenby RA, Silva AS, Gillies RJ, et al. (2009) Adaptive therapy. Cancer Res 69: 4894–4903. doi: 10.1158/0008-5472.CAN-08-3658
|
| [216] | Gatenby RA, Vincent TL (2003) An evolutionary model of carcinogenesis. Cancer Res 63: 6212–6220. |
| [217] |
Chan DA, Giaccia AJ (2007) Hypoxia, gene expression, and metastasis. Cancer Metastasis Rev 26: 333–339. doi: 10.1007/s10555-007-9063-1
|
| [218] |
Evans SM, Hahn SM, Magarelli DP, et al. (2001) Hypoxic heterogeneity in human tumors: EF5 binding, vasculature, necrosis, and proliferation. Am J Clin Oncol 24: 467–472. doi: 10.1097/00000421-200110000-00011
|
| [219] |
Martinive P, Defresne F, Quaghebeur E, et al. (2009) Impact of cyclic hypoxia on HIF-1alpha regulation in endothelial cells-new insights for anti-tumor treatments. FEBS J 276: 509–518. doi: 10.1111/j.1742-4658.2008.06798.x
|
| [220] |
Kim Y, Lin Q, Glazer PM, et al. (2009) Hypoxic tumor microenvironment and cancer cell differentiation. Curr Mol Med 9: 425–434. doi: 10.2174/156652409788167113
|
| [221] |
Harris AL (2002) Hypoxia-a key regulatory factor in tumour growth. Nat Rev Cancer 2: 38–47. doi: 10.1038/nrc704
|
| [222] |
Subarsky P, Hill RP (2003) The hypoxic tumour microenvironment and metastatic progression. Clin Exp Metastasis 20: 237–250. doi: 10.1023/A:1022939318102
|
| [223] |
Pouyssegur J, Dayan F, Mazure NM (2006) Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 441: 437–443. doi: 10.1038/nature04871
|
| [224] |
Semenza GL (2003) Targeting HIF-1 for cancer therapy. Nat Rev Cancer 3: 721–732. doi: 10.1038/nrc1187
|
| [225] | Struckmann K, Mertz K, Steu S, et al. (2008) pVHL co-ordinately regulates CXCR4/CXCL12 and MMP2/MMP9 expression in human clear-cell renal cell carcinoma. J Pathol 214: 464–471. |
| [226] |
Keith B, Simon MC (2007) Hypoxia-inducible factors, stem cells, and cancer. Cell 129: 465–472. doi: 10.1016/j.cell.2007.04.019
|
| [227] |
Bos R, Zhong H, Hanrahan CF, et al. (2001) Levels of hypoxia-inducible factor-1 alpha during breast carcinogenesis. J Natl Cancer Inst 93: 309–314. doi: 10.1093/jnci/93.4.309
|
| [228] |
Semenza GL (2003) Angiogenesis in ischemic and neoplastic disorders. Annu Rev Med 54: 17–28. doi: 10.1146/annurev.med.54.101601.152418
|
| [229] |
Serrati S, Margheri F, Fibbi G, et al. (2008) Endothelial cells and normal breast epithelial cells enhance invasion of breast carcinoma cells by CXCR-4-dependent up-regulation of urokinase-type plasminogen activator receptor (uPAR, CD87) expression. J Pathol 214: 545–554. doi: 10.1002/path.2309
|
| [230] | Jankowski K, Kucia M, Wysoczynski M, et al. (2003) Both hepatocyte growth factor (HGF) and stromal-derived factor-1 regulate the metastatic behavior of human rhabdomyosarcoma cells, but only HGF enhances their resistance to radiochemotherapy. Cancer Res 63: 7926–7935. |
| [231] |
Libura J, Drukala J, Majka M, et al. (2002) CXCR4-SDF-1 signaling is active in rhabdomyosarcoma cells and regulates locomotion, chemotaxis, and adhesion. Blood 100: 2597–2606. doi: 10.1182/blood-2002-01-0031
|
| [232] |
Maeda S, Shinchi H, Kurahara H, et al. (2008) CD133 expression is correlated with lymph node metastasis and vascular endothelial growth factor-C expression in pancreatic cancer. Br J Cancer 98: 1389–1397. doi: 10.1038/sj.bjc.6604307
|
| [233] |
Brabletz T, Jung A, Spaderna S, et al. (2005) Opinion: Migrating cancer stem cells-an integrated concept of malignant tumour progression. Nat Rev Cancer 5: 744–749. doi: 10.1038/nrc1694
|
| [234] |
Kang Y, Massague J (2004) Epithelial-mesenchymal transitions: Twist in development and metastasis. Cell 118: 277–279. doi: 10.1016/j.cell.2004.07.011
|
| [235] |
Pomerance M, Quillard J, Chantoux F, et al. (2006) High-level expression, activation, and subcellular localization of p38-MAP kinase in thyroid neoplasms. J Pathol 209: 298–306. doi: 10.1002/path.1975
|
| [236] |
Murphy PM (2001) Chemokines and the molecular basis of cancer metastasis. N Engl J Med 345: 833–835. doi: 10.1056/NEJM200109133451113
|
| [237] |
Stainier D (2006) No stem cell is an islet (yet). N Engl J Med 354: 521–523. doi: 10.1056/NEJMcibr055048
|
| [238] |
Zlotnik A (2008) New insights on the role of CXCR4 in cancer metastasis. J Pathol 215: 211–213. doi: 10.1002/path.2350
|
| [239] |
DeNardo DG, Johansson M, Coussens LM (2008) Immune cells as mediators of solid tumor metastasis. Cancer Metastasis Rev 27: 11–18. doi: 10.1007/s10555-007-9100-0
|
| [240] |
Hiratsuka S, Watanabe A, Sakurai Y, et al. (2008) The S100A8-serum amyloid A3-TLR4 paracrine cascade establishes a pre-metastatic phase. Nat Cell Biol 10: 1349–1355. doi: 10.1038/ncb1794
|
| [241] |
Bornstein SR, Hornsby PJ (2005) What can we learn from gene expression profiling for adrenal tumor management? J Clin Endocrinol Metab 90: 1900–1902. doi: 10.1210/jc.2005-0065
|
| [242] |
Garber ME, Troyanskaya OG, Schluens K, et al. (2001) Diversity of gene expression in adenocarcinoma of the lung. Proc Natl Acad Sci U S A 98: 13784–13789. doi: 10.1073/pnas.241500798
|
| [243] |
Steeg PS (2005) New insights into the tumor metastatic process revealed by gene expression profiling. Am J Pathol 166: 1291–1294. doi: 10.1016/S0002-9440(10)62348-6
|
| [244] |
van de Rijn M, Rubin BP (2002) Gene expression studies on soft tissue tumors. Am J Pathol 161: 1531–1534. doi: 10.1016/S0002-9440(10)64430-6
|
| [245] |
van't Veer LJ, Paik S, Hayes DF (2005) Gene expression profiling of breast cancer: A new tumor marker. J Clin Oncol 23: 1631–1635. doi: 10.1200/JCO.2005.12.005
|
| [246] | Vaupel P, Harrison L (2004) Tumor hypoxia: Causative factors, compensatory mechanisms, and cellular response. Oncologist 5: 4–9. |
| [247] |
Pennacchietti S, Michieli P, Galluzzo M, et al. (2003) Hypoxia promotes invasive growth by transcriptional activation of the met protooncogene. Cancer Cell 3: 347–361. doi: 10.1016/S1535-6108(03)00085-0
|
| [248] |
Kulshreshtha R, Davuluri RV, Calin GA, et al. (2008) A microRNA component of the hypoxic response. Cell Death Differ 15: 667–671. doi: 10.1038/sj.cdd.4402310
|
| [249] |
Ouatas T, Salerno M, Palmieri D, et al. (2003) Basic and translational advances in cancer metastasis: Nm23. J Bioenerg Biomembr 35: 73–79. doi: 10.1023/A:1023497924277
|
| [250] |
Salerno M, Ouatas T, Palmieri D, et al. (2003) Inhibition of signal transduction by the nm23 metastasis suppressor: Possible mechanisms. Clin Exp Metastasis 20: 3–10. doi: 10.1023/A:1022578000022
|
| [251] |
Steeg PS (2003) Metastasis suppressors alter the signal transduction of cancer cells. Nat Rev Cancer 3: 55–63. doi: 10.1038/nrc967
|
| [252] |
Steeg PS, Ouatas T, Halverson D, et al. (2003) Metastasis suppressor genes: Basic biology and potential clinical use. Clin Breast Cancer 4: 51–62. doi: 10.3816/CBC.2003.n.012
|
| [253] |
Wulfkuhle JD, Paweletz CP, Steeg PS, et al. (2003) Proteomic approaches to the diagnosis, treatment, and monitoring of cancer. Adv Exp Med Biol 532: 59–68. doi: 10.1007/978-1-4615-0081-0_7
|
| [254] |
Guo Y, Feng Y, Trivedi NS, et al. (2011) Medusa structure of the gene regulatory network: Dominance of transcription factors in cancer subtype classification. Exp Biol Med 236: 628–636. doi: 10.1258/ebm.2011.010324
|
| [255] |
Guo Z, Wu F, Asplund A, et al. (2001) Analysis of intratumoral heterogeneity of chromosome 3p deletions and genetic evidence of polyclonal origin of cervical squamous carcinoma. Mod Pathol 14: 54–61. doi: 10.1038/modpathol.3880256
|
| [256] |
Nakamura T, Kuwai T, Kitadai Y, et al. (2007) Zonal heterogeneity for gene expression in human pancreatic carcinoma. Cancer Res 67: 7597–7604. doi: 10.1158/0008-5472.CAN-07-0874
|
| [257] |
Shipitsin M, Campbell LL, Argani P, et al. (2007) Molecular definition of breast tumor heterogeneity. Cancer Cell 11: 259–273. doi: 10.1016/j.ccr.2007.01.013
|
| [258] |
Edelman EJ, Guinney J, Chi JT, et al. (2008) Modeling cancer progression via pathway dependencies. PLoS Comput Biol 4: e28. doi: 10.1371/journal.pcbi.0040028
|
| [259] | Califano J, van der Riet P, Westra W, et al. (1996) Genetic progression model for head and neck cancer: Implications for field cancerization. Cancer Res 56: 2488–2492. |
| [260] |
Parsons DW, Jones S, Zhang X, et al. (2008) An integrated genomic analysis of human glioblastoma multiforme. Science 321: 1807–1812. doi: 10.1126/science.1164382
|
| [261] |
Wood LD, Parsons DW, Jones S, et al. (2007) The genomic landscapes of human breast and colorectal cancers. Science 318: 1108–1113. doi: 10.1126/science.1145720
|
| [262] |
Siegmund KD, Marjoram P, Woo YJ, et al. (2009) Inferring clonal expansion and cancer stem cell dynamics from DNA methylation patterns in colorectal cancers. Proc Natl Acad Sci U S A 106: 4828–4833. doi: 10.1073/pnas.0810276106
|
| [263] |
Campbell PJ, Pleasance ED, Stephens PJ, et al. (2008) Subclonal phylogenetic structures in cancer revealed by ultra-deep sequencing. Proc Natl Acad Sci U S A 105: 13081–13086. doi: 10.1073/pnas.0801523105
|
| [264] |
Mullighan CG, Phillips LA, Su X, et al. (2008) Genomic analysis of the clonal origins of relapsed acute lymphoblastic leukemia. Science 322: 1377–1380. doi: 10.1126/science.1164266
|
| [265] |
Panzer-Grumayer ER, Cazzaniga G, van der Velden VH, et al. (2005) Immunogenotype changes prevail in relapses of young children with TEL-AML1-positive acute lymphoblastic leukemia and derive mainly from clonal selection. Clin Cancer Res 11: 7720–7727. doi: 10.1158/1078-0432.CCR-05-1239
|
| [266] |
Tomlinson IP, Lambros MB, Roylance RR (2002) Loss of heterozygosity analysis: Practically and conceptually flawed? Genes Chromosomes Cancer 34: 349–353. doi: 10.1002/gcc.10085
|
| [267] |
Park SY, Gonen M, Kim HJ, et al. (2010) Cellular and genetic diversity in the progression of in situ human breast carcinomas to an invasive phenotype. J Clin Invest 120: 636–644. doi: 10.1172/JCI40724
|
| [268] |
Diaz-Cano S (1996) PCR-based alternative for diagnosis of immunoglobulin heavy chain gene rearrangement: Principles, practice, and polemics. Diagn Mol Pathol 5: 3–9. doi: 10.1097/00019606-199603000-00002
|
| [269] |
Brady SP, Magro CM, Diaz-Cano SJ, et al. (1999) Analysis of clonality of atypical cutaneous lymphoid infiltrates associated with drug therapy by PCR/DGGE. Hum Pathol 30: 130–136. doi: 10.1016/S0046-8177(99)90266-6
|
| [270] | Teixeira MR, Pandis N, Bardi G, et al. (1996) Karyotypic comparisons of multiple tumorous and macroscopically normal surrounding tissue samples from patients with breast cancer. Cancer Res 56: 855–859. |
| [271] |
Teixeira MR, Pandis N, Bardi G, et al. (1995) Clonal heterogeneity in breast cancer: Karyotypic comparisons of multiple intra- and extra-tumorous samples from 3 patients. Int J Cancer 63: 63–68. doi: 10.1002/ijc.2910630113
|
| [272] | Giaretti W, Monaco R, Pujic N, et al. (1996) Intratumor heterogeneity of K-ras2 mutations in colorectal adenocarcinomas: Association with degree of DNA aneuploidy. Am J Pathol 149: 237–245. |
| [273] | Coons SW, Johnson PC, Shapiro JR (1995) Cytogenetic and flow cytometry DNA analysis of regional heterogeneity in a low grade human glioma. Cancer Res 55: 1569–1577. |
| [274] |
Kallioniemi OP (1988) Comparison of fresh and paraffin-embedded tissue as starting material for DNA flow cytometry and evaluation of intratumor heterogeneity. Cytometry 9: 164–169. doi: 10.1002/cyto.990090211
|
| [275] |
Lage JM, Leamon JH, Pejovic T, et al. (2003) Whole genome analysis of genetic alterations in small DNA samples using hyperbranched strand displacement amplification and array-CGH. Genome Res 13: 294–307. doi: 10.1101/gr.377203
|
| [276] | Leith JT, Michelson S, Faulkner LE, et al. (1987) Growth properties of artificial heterogeneous human colon tumors. Cancer Res 47: 1045–1051. |
| [277] |
Farber L, Efrati E, Elkin H, et al. (2011) Molecular morphometric analysis shows relative intra-tumoural homogeneity for KRAS mutations in colorectal cancer. Virchows Arch 459: 487–493. doi: 10.1007/s00428-011-1158-y
|
| [278] |
Kuwai T, Nakamura T, Kim SJ, et al. (2008) Intratumoral heterogeneity for expression of tyrosine kinase growth factor receptors in human colon cancer surgical specimens and orthotopic tumors. Am J Pathol 172: 358–366. doi: 10.2353/ajpath.2008.070625
|
| [279] |
Inda MM, Bonavia R, Mukasa A, et al. (2010) Tumor heterogeneity is an active process maintained by a mutant EGFR-induced cytokine circuit in glioblastoma. Genes Dev 24: 1731–1745. doi: 10.1101/gad.1890510
|
| [280] | Konishi N, Hiasa Y, Matsuda H, et al. (1995) Intratumor cellular heterogeneity and alterations in ras oncogene and p53 tumor suppressor gene in human prostate carcinoma. Am J Pathol 147: 1112–1122. |
| [281] | Maley CC, Reid BJ, Forrest S (2004) Cancer prevention strategies that address the evolutionary dynamics of neoplastic cells: Simulating benign cell boosters and selection for chemosensitivity. Cancer Epidemiol Biomarkers Prev 13: 1375–1384. |
| [282] | Cottu PH, Asselah J, Lae M, et al. (2008) Intratumoral heterogeneity of HER2/neu expression and its consequences for the management of advanced breast cancer. Ann Oncol 19: 595–597. |
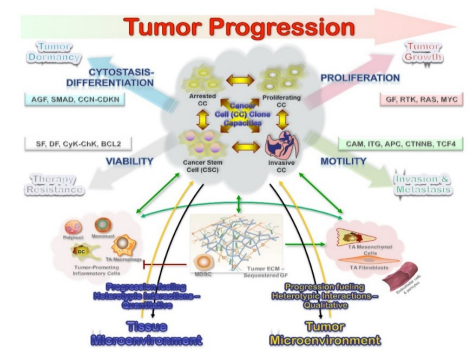
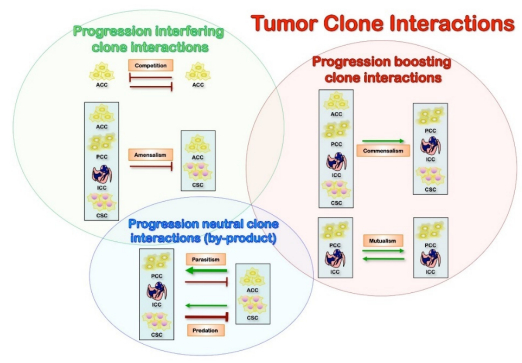
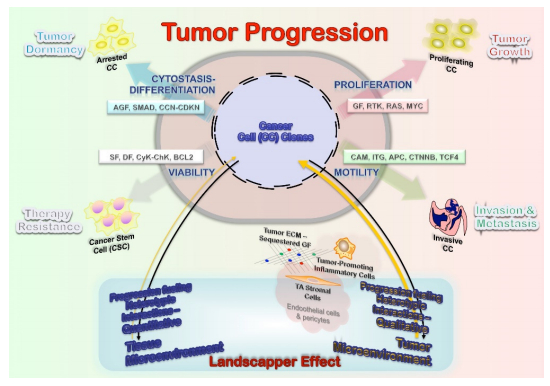
Figures(3)

Fatima Al-Hashimi, Salvador J. Diaz-Cano. Multi-target analysis of neoplasms for the evaluation of tumor progression: stochastic approach of biologic processes[J]. AIMS Molecular Science, 2018, 5(1): 14-62. doi: 10.3934/molsci.2018.1.14







 DownLoad:
DownLoad: