Citation: Jon Knight, Pat Dale, Patrick Dwyer, Sam Marx. A conceptual approach to integrate management of ecosystem service and disservice in coastal wetlands[J]. AIMS Environmental Science, 2017, 4(3): 431-442. doi: 10.3934/environsci.2017.3.431

| [1] |
Schmidt JP, Moore R, Alber M (2014) Integrating ecosystem services and local government finances into land use planning: a case study from coastal Georgia. Landscape Urban Plan 122: 56-67. doi: 10.1016/j.landurbplan.2013.11.008

|
| [2] |
Costanza R, de Groot R, Sutton P, et al. (2014) Changes in the global value of ecosystem services. Global Environ Chang 26: 152-158. doi: 10.1016/j.gloenvcha.2014.04.002
|
| [3] |
Rao NS, Ghermandi A, Portela R, et al. (2015) Global values of coastal ecosystem services: a spatial economic analysis of shoreline protection values. Ecosyst Serv 11: 95-105. doi: 10.1016/j.ecoser.2014.11.011
|
| [4] |
Barbier EB, Hacker SD, Kennedy C, et al. (2011) The value of estuarine and coastal ecosystem services. Ecol Monogr 81: 169-193. doi: 10.1890/10-1510.1
|
| [5] |
Rog SM, Clarke RH, Cook CN (2017) More than marine: revealing the critical importance of mangrove ecosystems for terrestrial vertebrates. Divers Distrib 23: 221-230. doi: 10.1111/ddi.12514
|
| [6] |
Gurran N (2008) The turning tide: amenity migration in coastal Australia. Int Plan Stud 13: 391-414. doi: 10.1080/13563470802519055
|
| [7] |
Dale PER, Knight JM, Dwyer PG (2014) Mangrove rehabilitation: a review focusing on ecological and institutional issues. Wetl Ecol Manag 22: 587-604. doi: 10.1007/s11273-014-9383-1
|
| [8] | Adam P (2002) Saltmarshes in a time of change. Environ Conserv 29: 39-61. |
| [9] |
Saintilan N, Rogers K (2013) The significance and vulnerability of Australian saltmarshes: implications for management in a changing climate. Mar Freshwater Res 64: 66-79. doi: 10.1071/MF12212
|
| [10] |
Lyytimäki J, Sipilä M (2009) Hopping on one leg-the challenge of ecosystem disservices for urban green management. Urban For Urban Gree 8: 309-315. doi: 10.1016/j.ufug.2009.09.003
|
| [11] |
Friess DA (2016) Ecosystem services and disservices of mangrove forests: insights from historical colonial observations. Forests 7: 183. doi: 10.3390/f7090183
|
| [12] |
Lyytimäki J (2015) Ecosystem disservices: embrace the catchword. Ecosyst Serv 12: 136. doi: 10.1016/j.ecoser.2014.11.008
|
| [13] | Lyytimäki J, Petersen LK, Normander B, et al. (2008) Nature as a nuisance? Ecosystem services and disservices to urban lifestyle. Environ Sci 5: 161-172. |
| [14] |
Fisher B, Turner RK, Morling P (2009) Defining and classifying ecosystem services for decision making. Ecol Econ 68: 643-653. doi: 10.1016/j.ecolecon.2008.09.014
|
| [15] | Dwyer PG, Knight JM, Dale PER (2016) Planning development to reduce mosquito hazard in coastal peri-urban areas: case studies in NSW, Australia. In: Maheshwari B, Singh VP, Thoradeniya B, editors. Balanced urban development: options and strategies for liveable cities. Switzerland: Springer International Publishing AG. pp. 555-574. |
| [16] |
Knight JM (2011) A model of mosquito–mangrove basin ecosystems with implications for management. Ecosystems 14: 1382-1395. doi: 10.1007/s10021-011-9487-x
|
| [17] | Webb C (2013) Managing mosquitos in coastal wetlands. WET eBook workbook for managing urban wetlands in Australia Sydney Olympic Park. |
| [18] | Provost MW (1977) Source reduction in salt-marsh mosquito control: past and future. Mosquito News 37: 689-698. |
| [19] |
Rey J, Walton W, Wolfe R, et al. (2012) North American wetlands and mosquito control. Inter J Env Res Pub Heal 9: 4537-4605. doi: 10.3390/ijerph9124537
|
| [20] |
Carlson DB (2006) Source reduction in Florida's salt marshes: management to reduce pesticide use and enhance the resource. J Am Mosquito Contr 22: 534-537. doi: 10.2987/8756-971X(2006)22[534:SRIFSM]2.0.CO;2
|
| [21] | Wolfe RJ (1996) Effects of open marsh water management on selected tidal marsh resources: a review. J Am Mosquito Contr 12: 701-712. |
| [22] |
Dale PER (2008) Assessing impacts of habitat modification on a subtropical salt marsh: 20 years of monitoring. Wetl Ecol Manag 16: 77-87. doi: 10.1007/s11273-007-9058-2
|
| [23] | Hulsman K, Dale PER, Kay BH (1989) The runnelling method of habitat modification: an environment focused tool for salt marsh management. J Am Mosquito Contr 5: 226-234. |
| [24] |
Dale P, Knight J (2012) Managing mosquitoes without destroying wetlands: an eastern Australian approach. Wetl Ecol Manag 20: 233-242. doi: 10.1007/s11273-012-9262-6
|
| [25] |
Knight JM, Dale PER, Spencer J, et al. (2009) Exploring LiDAR data for mapping the micro-topography and tidal hydro-dynamics of mangrove systems: an example from southeast Queensland, Australia. Estuar Coast Shelf S 85: 593-600. doi: 10.1016/j.ecss.2009.10.002
|
| [26] |
Dale PER, Dale MB, Dowe DL, et al. (2010) A conceptual model for integrating physical geography research and coastal wetland management, with an Australian example. Prog Phys Geog 34: 605-624. doi: 10.1177/0309133310369617
|
| [27] | Harley D, Ritchie S, Bain C, et al. (2005) Risks for ross river virus disease in tropical Australia. Int J Epidemiol 34: 548-555. |
| [28] |
Knight JM, Griffin L, Dale PER, et al. (2013) Short-term dissolved oxygen patterns in sub-tropical mangroves. Estuar Coast Shelf S 131: 290-296. doi: 10.1016/j.ecss.2013.06.024
|
| [29] | Meynecke JO, Lee SY, Lofthouse J, et al. (2013) Fish In, Mozzies Out: Final Report to the NSW Recreational Fishing Trust. Griffith University, Brisbane, Queensland Australia. (ISBN 978-1-922216-25-0). |
| [30] |
Dale PE, Knight JM, Griffin L, et al. (2014) Multi-agency perspectives on managing mangrove wetlands and the mosquitoes they produce. J Am Mosquito Contr 30: 106-115. doi: 10.2987/13-6393.1
|
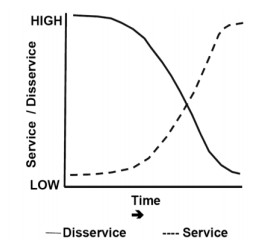
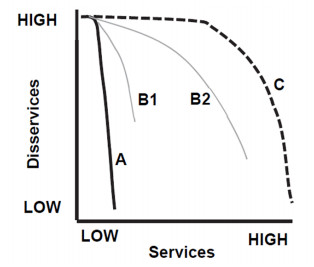
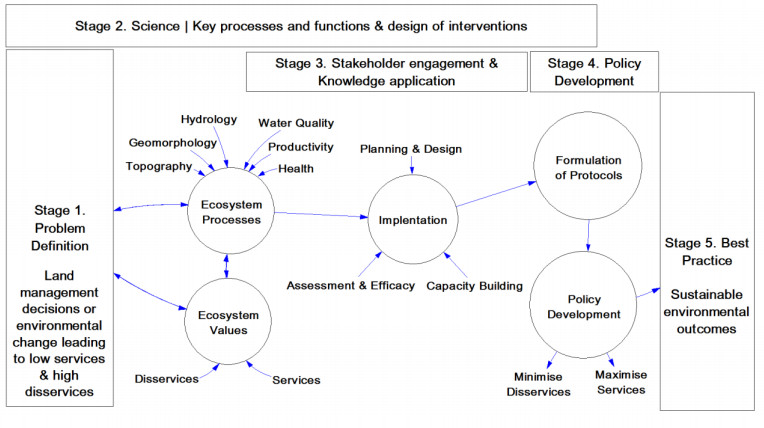
Figures(3)

Jon Knight, Pat Dale, Patrick Dwyer, Sam Marx. A conceptual approach to integrate management of ecosystem service and disservice in coastal wetlands[J]. AIMS Environmental Science, 2017, 4(3): 431-442. doi: 10.3934/environsci.2017.3.431







 DownLoad:
DownLoad: