Citation: Stephen H. Kasper, Ryan Hart, Magnus Bergkvist, Rabi A. Musah, Nathaniel C. Cady. Zein nanocapsules as a tool for surface passivation, drug delivery and biofilm prevention[J]. AIMS Microbiology, 2016, 2(4): 422-433. doi: 10.3934/microbiol.2016.4.422

| [1] |
Kolenbrander PE, Palmer RJ, Rickard AH, et al. (2006) Bacterial interactions and successions during plaque development. Periodontol 2000 42: 47–79. doi: 10.1111/j.1600-0757.2006.00187.x

|
| [2] |
Marsh PD, Moter A, Devine DA (2011) Dental plaque biofilms: communities, conflict and control. Periodontol 2000 55: 16–35. doi: 10.1111/j.1600-0757.2009.00339.x
|
| [3] |
Costerton JW (1999) Bacterial biofilms: a common cause of persistent infections. Science 284: 1318–1322. doi: 10.1126/science.284.5418.1318
|
| [4] |
Costalonga M, Herzberg MC (2014) The oral microbiome and the immunobiology of periodontal disease and caries. Immunol Lett 162: 22–38. doi: 10.1016/j.imlet.2014.08.017
|
| [5] |
Kumar PS (2013) Oral microbiota and systemic disease. Anaerobe 24: 90–93. doi: 10.1016/j.anaerobe.2013.09.010
|
| [6] |
Han YW, Wang X (2013) Mobile microbiome: oral bacteria in extra-oral infections and inflammation. J Dent Res 92: 485–91. doi: 10.1177/0022034513487559
|
| [7] |
Scannapieco FA, Cantos A (2016) Oral inflammation and infection, and chronic medical diseases: implications for the elderly. Periodontol 2000 72: 153–175. doi: 10.1111/prd.12129
|
| [8] |
Marsh PD (2010) Controlling the oral biofilm with antimicrobials. J Dent 38: S11–S15. doi: 10.1016/S0300-5712(10)70005-1
|
| [9] |
Huang R, Li M, Gregory RL (2011) Bacterial interactions in dental biofilm. Virulence 2: 435–444. doi: 10.4161/viru.2.5.16140
|
| [10] |
Kasper SH, Samarian D, Jadhav AP, et al. (2014) S-aryl-L-cysteine sulphoxides and related organosulphur compounds alter oral biofilm development and AI-2-based cell-cell communication. J Appl Microbiol 117: 1472–1486. doi: 10.1111/jam.12616
|
| [11] |
Liu C, Worthington RJ, Melander C, et al. (2011) A new small molecule specifically inhibits the cariogenic bacterium Streptococcus mutans in multispecies biofilms. Antimicrob Agents Chemother 55: 2679–2687. doi: 10.1128/AAC.01496-10
|
| [12] |
He Z, Wang Q, Hu Y, et al. (2012) Use of the quorum sensing inhibitor furanone C-30 to interfere with biofilm formation by Streptococcus mutans and its luxS mutant strain. Int J Antimicrob Agents 40: 30–35. doi: 10.1016/j.ijantimicag.2012.03.016
|
| [13] |
Kunze B, Reck M, Dötsch A, et al. (2010) Damage of Streptococcus mutans biofilms by carolacton, a secondary metabolite from the myxobacterium Sorangium cellulosum. BMC Microbiol 10: 199. doi: 10.1186/1471-2180-10-199
|
| [14] |
Eguchi Y, Kubo N, Matsunaga H, et al. (2011) Development of an antivirulence drug against Streptococcus mutans: repression of biofilm formation, acid tolerance, and competence by a histidine kinase inhibitor, walkmycin C. Antimicrob Agents Chemother 55: 1475–1484. doi: 10.1128/AAC.01646-10
|
| [15] |
Jones MN, Francis SE, Hutchinson FJ, et al. (1993) Targeting and delivery of bactericide to adsorbed oral bacteria by use of proteoliposomes. Biochim Biophys Acta 1147: 251–261. doi: 10.1016/0005-2736(93)90010-W
|
| [16] |
Robinson AM, Creeth JE, Jones MN (2000) The use of immunoliposomes for specific delivery of antimicrobial agents to oral bacteria immobilized on polystyrene. J Biomater Sci 11: 1381–1393. doi: 10.1163/156856200744408
|
| [17] |
Jones MN (2005) Use of liposomes to deliver bactericides to bacterial biofilms. Methods Enzymol 391: 211–228. doi: 10.1016/S0076-6879(05)91013-6
|
| [18] |
Chen F, Liu XM, Rice KC, et al. (2009) Tooth-binding micelles for dental caries prevention. Antimicrob Agents Chemother 53: 4898–4902. doi: 10.1128/AAC.00387-09
|
| [19] |
Chen F, Jia Z, Rice KC, et al. (2013) The development of dentotropic micelles with biodegradable tooth-binding moieties. Pharm Res 30: 2808–2817. doi: 10.1007/s11095-013-1105-5
|
| [20] | Chen F, Rice KC, Liu XM, et al. (2010) Triclosan-loaded tooth-binding micelles for prevention and treatment of dental biofilm, Pharm Res 27: 2356–2364. |
| [21] |
Horev B, Klein MI, Hwang G, et al. (2015) pH-activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano 9: 2390–2404. doi: 10.1021/nn507170s
|
| [22] |
Radovic-Moreno AF, Lu TK, Puscasu VA, et al. (2012) Surface charge-switching polymeric nanoparticles for bacterial cell wall-targeted delivery of antibiotics. ACS Nano 6: 4279–4287. doi: 10.1021/nn3008383
|
| [23] |
Hannig M, Hannig C (2010) Nanomaterials in preventive dentistry. Nat Nanotechnol 5: 565–569. doi: 10.1038/nnano.2010.83
|
| [24] |
Nie B, Chen T, Liang M, et al. (2010) The use of nanoparticles to control oral biofilm formation. J Dent Res 89: 1175–1186. doi: 10.1177/0022034510377794
|
| [25] | Murdan S (2005) Formulation and characterisation of zein microspheres as delivery vehicles. J Drug Deliv Sci Technol 15: 267–272. |
| [26] |
Shukla R, Cheryan M (2001) Zein: the industrial protein from corn. Ind Crops Prod 13: 171–192. doi: 10.1016/S0926-6690(00)00064-9
|
| [27] |
Elzoghby AO, Samy WM, Elgindy NA (2012) Protein-based nanocarriers as promising drug and gene delivery systems. J Control Release 161: 38–49. doi: 10.1016/j.jconrel.2012.04.036
|
| [28] |
Regier MC, Taylor JD, Borcyk T, et al. (2012) Fabrication and characterization of DNA-loaded zein nanospheres. J Nanobiotechnology 10: 44. doi: 10.1186/1477-3155-10-44
|
| [29] |
Wang HJ, Lin ZX, Liu XM, et al. (2005) Heparin-loaded zein microsphere film and hemocompatibility. J Control Release 105: 120–131. doi: 10.1016/j.jconrel.2005.03.014
|
| [30] |
Torres-Giner S, Martinez-Abad A, Ocio MJ, et al. (2010) Stabilization of a nutraceutical omega-3 fatty acid by encapsulation in ultrathin electrosprayed zein prolamine. J Food Sci 75: N69–N79. doi: 10.1111/j.1750-3841.2010.01678.x
|
| [31] |
Luo Y, Teng Z, Wang Q (2012) Development of zein nanoparticles coated with carboxymethyl chitosan for encapsulation and controlled release of vitamin D3. J Agric Food Chem 60: 836–843. doi: 10.1021/jf204194z
|
| [32] |
Fu JX, Wang HJ, Zhou YQ, et al. (2009) Antibacterial activity of ciprofloxacin-loaded zein microsphere films. Mater Sci Eng C 29: 1161–1166. doi: 10.1016/j.msec.2008.09.031
|
| [33] |
Panchapakesan C, Sozer N, Dogan H, et al. (2012) Effect of different fractions of zein on the mechanical and phase properties of zein films at nano-scale. J Cereal Sci 55: 174–182. doi: 10.1016/j.jcs.2011.11.004
|
| [34] |
Cady NC, McKean KA, Behnke J, et al. (2012) Inhibition of biofilm formation, quorum sensing and infection in Pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS One 7: e38492. doi: 10.1371/journal.pone.0038492
|
| [35] |
Welin J, Wilkins JC, Beighton D, et al. (2004) Protein expression by Streptococcus mutans during initial stage of biofilm formation. Appl Environ Microbiol 70: 3736–3741. doi: 10.1128/AEM.70.6.3736-3741.2004
|
| [36] | Koo H, Hayacibara MF, Schobel BD, et al. (2003) Inhibition of Streptococcus mutans biofilm accumulation and polysaccharide production by apigenin and tt-farnesol, J Antimicrob Chemother 52: 782–789. |
| [37] |
Merritt J, Qi F, Goodman SD, et al. (2003) Mutation of luxS affects biofilm formation in Streptococcus mutans. Infect Immun 71: 1972–1979. doi: 10.1128/IAI.71.4.1972-1979.2003
|
| [38] |
Landini P, Antoniani D, Burgess JG, et al. (2010) Molecular mechanisms of compounds affecting bacterial biofilm formation and dispersal. Appl Microbiol Biotechnol 86: 813–823. doi: 10.1007/s00253-010-2468-8
|
| [39] |
Whitchurch CB, Tolker-Nielsen T, Ragas PC, et al. (2002) Extracellular DNA required for bacterial biofilm formation. Science 295: 1487–1487. doi: 10.1126/science.295.5559.1487
|
| [40] |
Perry JA, Cvitkovitch DG, Levesque CM (2009) Cell death in Streptococcus mutans biofilms: a link between CSP and extracellular DNA. FEMS Microbiol Lett 299: 261–266. doi: 10.1111/j.1574-6968.2009.01758.x
|
| [41] | Allaker RP, Memarzadeh K (2014) Nanoparticles and the control of oral infections. Int J Antimicrob Agents 43: 95–104. |
| [42] |
Melo MAS, Guedes SFF, Xu HHK, et al. (2013) Nanotechnology-based restorative materials for dental caries management. Trends Biotechnol 31: 459–467. doi: 10.1016/j.tibtech.2013.05.010
|
| [43] |
Benoit DSW, Koo H (2016) Targeted, triggered drug delivery to tumor and biofilm microenvironments. Nanomedicine 11: 873–879. doi: 10.2217/nnm-2016-0014
|
| [44] |
Forier K, Raemdonck K, De Smedt SC, et al. (2014) Lipid and polymer nanoparticles for drug delivery to bacterial biofilms. J Control Release 190: 607–623. doi: 10.1016/j.jconrel.2014.03.055
|
| [45] |
Torres-Giner S, Ocio MJ, Lagaron JM (2009) Novel antimicrobial ultrathin structures of zein/chitosan blends obtained by electrospinning. Carbohydr Polym 77: 261–266. doi: 10.1016/j.carbpol.2008.12.035
|
| [46] |
Del Nobile MA, Conte A, Incoronato AL, et al. (2008) Antimicrobial efficacy and release kinetics of thymol from zein films. J Food Eng 89: 57–63. doi: 10.1016/j.jfoodeng.2008.04.004
|
| [47] |
Dawson PL, Hirt DE, Rieck JR, et al. (2003) Nisin release from films is affected by both protein type and film-forming method. Food Res Int 36: 959–968. doi: 10.1016/S0963-9969(03)00116-9
|
| [48] |
Sousa FFO, Luzardo-Alvarez A, Pérez-Estévéz A, et al. (2010) Development of a novel AMX-loaded PLGA/zein microsphere for root canal disinfection. Biomed Mater 5: 055008. doi: 10.1088/1748-6041/5/5/055008
|
| [49] |
de Sousa FO, Blanco-Méndez J, Pérez-Estévez A, et al. (2012) Effect of zein on biodegradable inserts for the delivery of tetracycline within periodontal pockets. J Biomater Appl 27: 187–200. doi: 10.1177/0885328211398968
|
| [50] |
Yun J, Fan X, Li X, et al. (2015) Natural surface coating to inactivate Salmonella enterica serovar Typhimurium and maintain quality of cherry tomatoes. Int J Food Microbiol 193: 59–67. doi: 10.1016/j.ijfoodmicro.2014.10.013
|
| [51] |
Lai LF, Guo HX (2011) Preparation of new 5-fluorouracil-loaded zein nanoparticles for liver targeting. Int J Pharm 404: 317–323. doi: 10.1016/j.ijpharm.2010.11.025
|
| [52] |
Muthuselvi L, Dhathathreyan A (2006) Simple coacervates of zein to encapsulate Gitoxin. Colloids Surf B Biointerfaces 51: 39–43. doi: 10.1016/j.colsurfb.2006.05.012
|
| [53] |
Chen L, Subirade M (2009) Elaboration and characterization of soy/zein protein microspheres for controlled nutraceutical delivery. Biomacromolecules 10: 3327–3334. doi: 10.1021/bm900989y
|
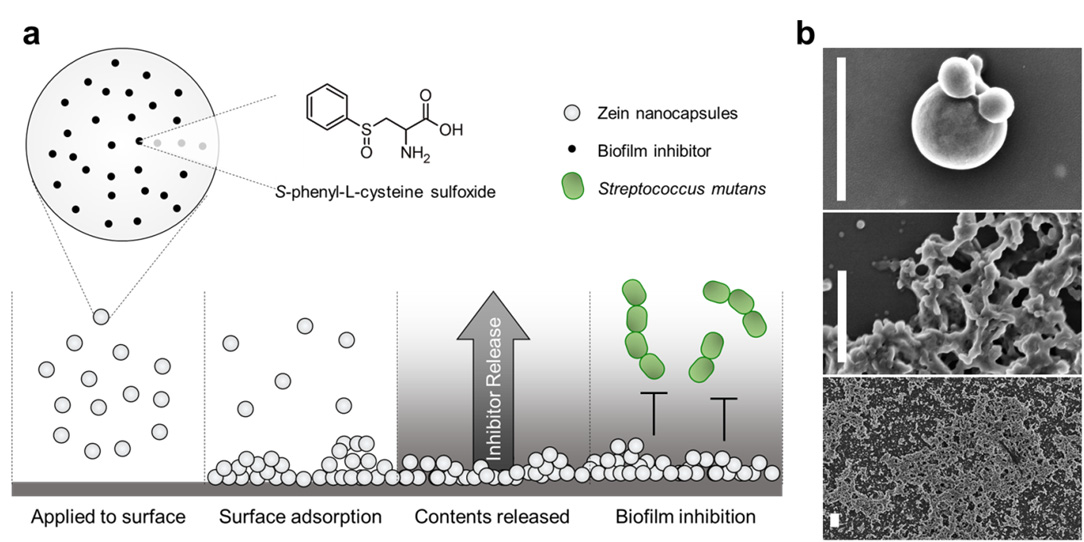
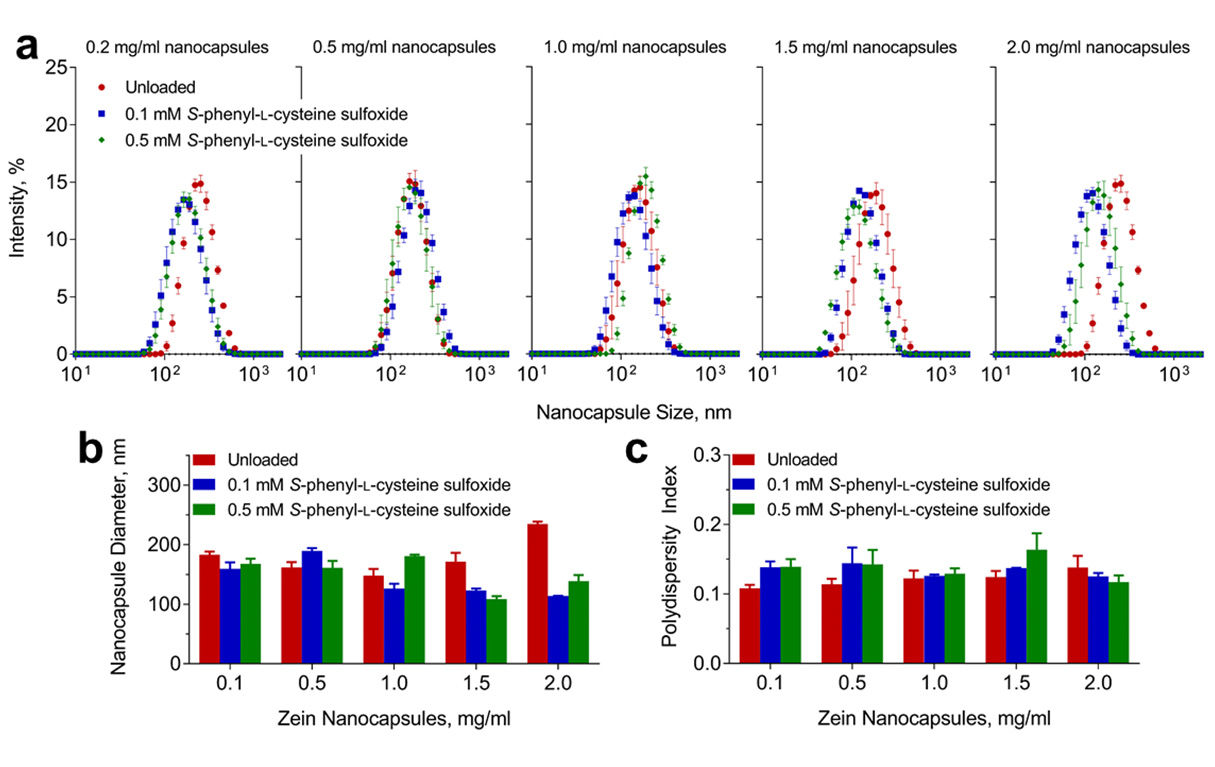
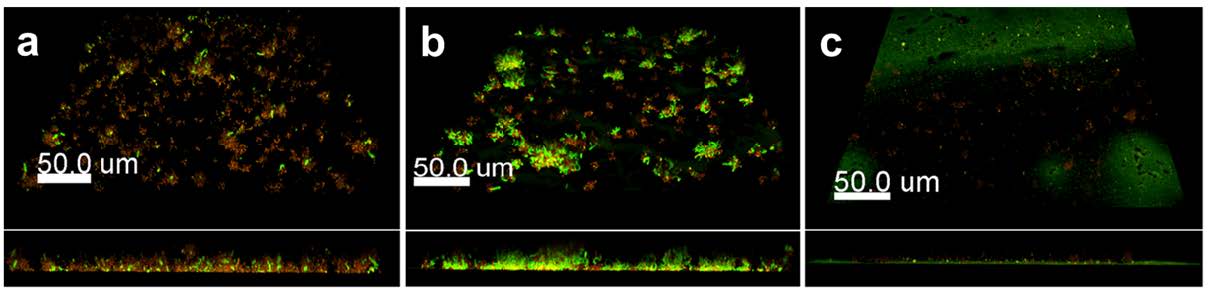
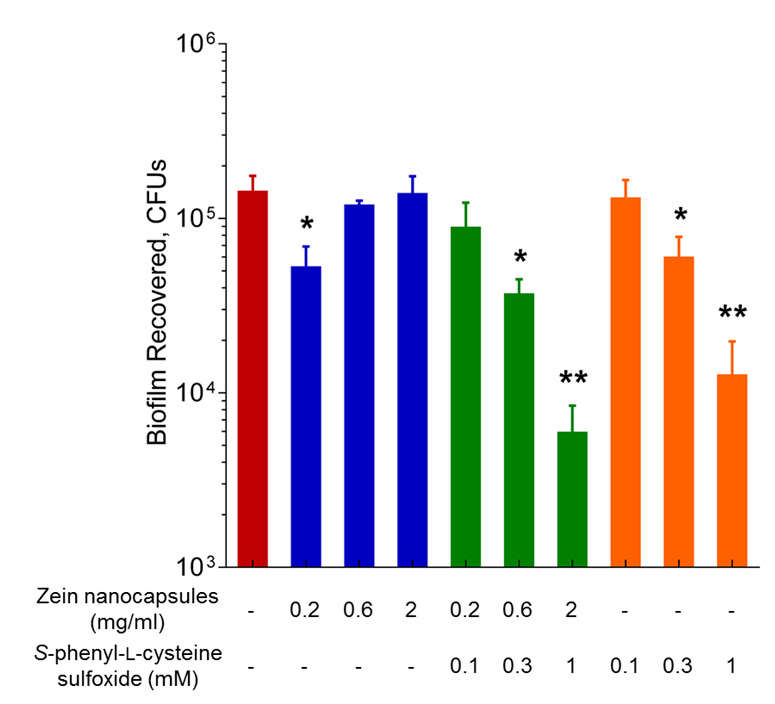
Figures(4)

Stephen H. Kasper, Ryan Hart, Magnus Bergkvist, Rabi A. Musah, Nathaniel C. Cady. Zein nanocapsules as a tool for surface passivation, drug delivery and biofilm prevention[J]. AIMS Microbiology, 2016, 2(4): 422-433. doi: 10.3934/microbiol.2016.4.422







 DownLoad:
DownLoad: