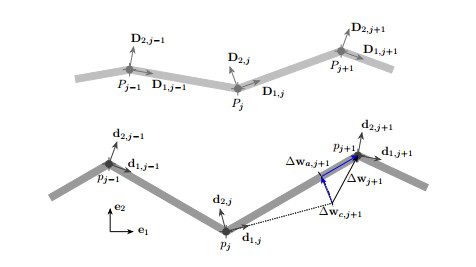
Mechanical metamaterials are most often assemblies of stocky beam elements connected through rigid connections, hinges, or flexural joints. The description of these materials through classical beam theories is challenging because of the wide variety of complex phenomena observed in the severe deformation regime mechanical metamaterials must undergo and because most classical beam theories can only be applied to elements with sufficiently high slenderness. In the spirit of Hencky, Turco et al. (2020) has recently formulated an intrinsically discrete nonlinear elastic model suitable for the design of mechanical metamaterials. The objective of this contribution was to present a numerical study of the nonlinear generalization of the Timoshenko beam that results from the asymptotic homogenization of the discrete model introduced by Turco et al. The present numerical study took into account several loading cases and elucidated the sensitivity of the homogenized continuum with respect to axial, bending, and shear stiffness parameters, as well as to load imperfections, in terms of mechanical behavior, including buckling onset and post-critical behavior. It was found that the predictions obtained with the homogenized model in the large deformation regime matched excellently with those of the discrete model proposed by Turco et al.
Citation: Jose Manuel Torres Espino, Emilio Barchiesi. Computational study of a homogenized nonlinear generalization of Timoshenko beam proposed by Turco et al.[J]. Networks and Heterogeneous Media, 2024, 19(3): 1133-1155. doi: 10.3934/nhm.2024050
Mechanical metamaterials are most often assemblies of stocky beam elements connected through rigid connections, hinges, or flexural joints. The description of these materials through classical beam theories is challenging because of the wide variety of complex phenomena observed in the severe deformation regime mechanical metamaterials must undergo and because most classical beam theories can only be applied to elements with sufficiently high slenderness. In the spirit of Hencky, Turco et al. (2020) has recently formulated an intrinsically discrete nonlinear elastic model suitable for the design of mechanical metamaterials. The objective of this contribution was to present a numerical study of the nonlinear generalization of the Timoshenko beam that results from the asymptotic homogenization of the discrete model introduced by Turco et al. The present numerical study took into account several loading cases and elucidated the sensitivity of the homogenized continuum with respect to axial, bending, and shear stiffness parameters, as well as to load imperfections, in terms of mechanical behavior, including buckling onset and post-critical behavior. It was found that the predictions obtained with the homogenized model in the large deformation regime matched excellently with those of the discrete model proposed by Turco et al.

| [1] |
J. Alibert, P. Seppecher, F. dell'Isola, Truss modular beams with deformation energy depending on higher displacement gradients, Math Mech Solids, 8 (2023), 51–73. https://doi.org/10.1177/1081286503008001658 doi: 10.1177/1081286503008001658

|
| [2] |
H. Altenbach, V. Eremeyev, On the constitutive equations of viscoelastic micropolar plates and shells of differential type, Math. Mech. Complex Syst., 3 (2015), 273–283. https://doi.org/10.2140/memocs.2015.3.273 doi: 10.2140/memocs.2015.3.273
|
| [3] | E. Barchiesi, Multi-scale and multi-physics: towards next-generation engineering materials, Continuum Mech. Thermodyn., 32 (2020), 541–554. |
| [4] |
E. Barchiesi, F. dell'Isola, A. M. Bersani, E. Turco, Equilibria determination of elastic articulated duoskelion beams in 2d via a riks-type algorithm, Int. J. Non Linear Mech, 128 (2021), 103628. https://doi.org/10.1016/j.ijnonlinmec.2020.103628 doi: 10.1016/j.ijnonlinmec.2020.103628
|
| [5] |
E. Barchiesi, F. dell'Isola, F. Hild, On the validation of homogenized modeling for bi-pantographic metamaterials via digital image correlation, Int J Solids Struct, 208 (2021), 49–62. https://doi.org/10.1016/j.ijsolstr.2020.09.036 doi: 10.1016/j.ijsolstr.2020.09.036
|
| [6] |
K. Barri, Q. Zhang, J. Kline, W. Lu, J. Luo, Z. Sun, et al., Multifunctional nanogenerator-integrated metamaterial concrete systems for smart civil infrastructure, Adv. Mater., 35 (2023), 2211027. https://doi.org/10.1002/adma.202211027 doi: 10.1002/adma.202211027
|
| [7] |
R. P. Bohara, S. Linforth, T. Nguyen, A. Ghazlan, T. Ngo, Anti-blast and -impact performances of auxetic structures: A review of structures, materials, methods, and fabrications, Eng Struct, 276 (2023), 115377. https://doi.org/10.1016/j.engstruct.2022.115377 doi: 10.1016/j.engstruct.2022.115377
|
| [8] |
C. Boutin, F. dell'Isola, Green's functions and integral representation of generalized continua: the case of orthogonal pantographic lattices, Z. Angew. Math. Phys., 72 (2021), 58. https://doi.org/10.1007/s00033-021-01480-3 doi: 10.1007/s00033-021-01480-3
|
| [9] |
M. S. Chaki, V. A. Eremeyev, A. K. Singh, Surface and interfacial anti-plane waves in micropolar solids with surface energy, Math Mech Solids, 26 (2021), 708–721. https://doi.org/10.1177/1081286520965646 doi: 10.1177/1081286520965646
|
| [10] |
F. Cornacchia, F. Fabbrocino, N. Fantuzzi, R. Luciano, R. Penna, Analytical solution of cross-and angle-ply nano plates with strain gradient theory for linear vibrations and buckling, Mech Adv. Mater. Struct, 28 (2021), 1201–1215. https://doi.org/10.1080/15376494.2019.1655613 doi: 10.1080/15376494.2019.1655613
|
| [11] |
H. Darban, R. Luciano, A. Caporale, F. Fabbrocino., Higher modes of buckling in shear deformable nanobeams, Int J Eng Sci, 154 (2020), 103338. https://doi.org/10.1016/j.ijengsci.2020.103338 doi: 10.1016/j.ijengsci.2020.103338
|
| [12] |
M. De Angelo, L. Placidi, N. Nejadsadeghi, A. Misra, Non-standard timoshenko beam model for chiral metamaterial: Identification of stiffness parameters, Mech Res Commun, 103 (2020), 103462. https://doi.org/10.1016/j.ijengsci.2020.103338 doi: 10.1016/j.ijengsci.2020.103338
|
| [13] |
I. Elishakoff, Who developed the so-called timoshenko beam theory?, Math Mech Solids, 25 (2020), 97–116. https://doi.org/10.1177/1081286519856931 doi: 10.1177/1081286519856931
|
| [14] | V. Eremeyev, Wojciech Pietraszkiewicz, Refined theories of plates and shells, ZAMM-Z Angew Math Me, 94 (2014), 5. |
| [15] |
V. A. Eremeyev, Two-and three-dimensional elastic networks with rigid junctions: modeling within the theory of micropolar shells and solids, Acta Mech, 230 (2019), 3875–3887. https://doi.org/10.1007/s00707-019-02527-3 doi: 10.1007/s00707-019-02527-3
|
| [16] |
V. A. Eremeyev, L. P. Lebedev, V. Konopińska-Zmysłowska, On solvability of initial boundary-value problems of micropolar elastic shells with rigid inclusions, Math Mech Solids, 27 (2022), 1800–1812. https://doi.org/10.1177/10812865211073149 doi: 10.1177/10812865211073149
|
| [17] |
V. A. Eremeyev, W. Pietraszkiewicz, Material symmetry group and constitutive equations of micropolar anisotropic elastic solids, Math Mech Solids, 21 (2016), 210–221. https://doi.org/10.1177/1081286515582862 doi: 10.1177/1081286515582862
|
| [18] |
V. A. Eremeyev, E. Turco, Enriched buckling for beam-lattice metamaterials, Mech Res Commun, 103 (2020), 103458. https://doi.org/10.1016/j.mechrescom.2019.103458 doi: 10.1016/j.mechrescom.2019.103458
|
| [19] |
F. Fabbrocino, M. Funari, F. Greco, P. Lonetti, R. Luciano, R. Penna, Dynamic crack growth based on moving mesh method, Compos Part B-eng, 174 (2019), 107053. https://doi.org/10.1016/j.compositesb.2019.107053 doi: 10.1016/j.compositesb.2019.107053
|
| [20] |
N. Feng, Y. Tie, S. Wang, J. Guo, A novel 3D bidirectional auxetic metamaterial with lantern-shape: Elasticity aspects and potential for load-bearing structure, Compos Struct, 321 (2023), 117221. https://doi.org/10.1016/j.compstruct.2023.117221 doi: 10.1016/j.compstruct.2023.117221
|
| [21] |
M. F. Funari, S. Spadea, F. Fabbrocino, R. Luciano, A moving interface finite element formulation to predict dynamic edge debonding in frp-strengthened concrete beams in service conditions, Fibers, 8 (2020), 42. https://doi.org/10.3390/fib8060042 doi: 10.3390/fib8060042
|
| [22] |
I. Giorgio, F. Hild, E. Gerami, F. dell'Isola, A. Misra, Experimental verification of 2D cosserat chirality with stretch-micro-rotation coupling in orthotropic metamaterials with granular motif, Mech Res Commun, 126 (2022), 104020. https://doi.org/10.1016/j.mechrescom.2022.104020 doi: 10.1016/j.mechrescom.2022.104020
|
| [23] |
I. Giorgio, V. Varano, F. dell'Isola, N. L. Rizzi, Two layers pantographs: A 2D continuum model accounting for the beams' offset and relative rotations as averages in SO(3) lie groups, Int J Solids Struct, 216 (2021), 43–58. https://doi.org/10.1016/j.ijsolstr.2021.01.018 doi: 10.1016/j.ijsolstr.2021.01.018
|
| [24] |
D. Han, X. Ren, Y. Zhang, X. Y. Zhang, X. G. Zhang, C. Luo, Y. M. Xie, Lightweight auxetic tamaterials: Design and characteristic study, Compos Struct, 293 (2022), 115706. https://doi.org/10.1016/j.compstruct.2022.115706 doi: 10.1016/j.compstruct.2022.115706
|
| [25] | H. Hencky, Über die angenäherte Lösung von Stabilitätsproblemen im Raum mittels der elastischen Gelenkkette, (Germany), Doctoral Thesis of W. Engelmann, Leipzig, 1921. |
| [26] |
F. Hild, A. Misra, F. dell'Isola, Multiscale DIC applied to pantographic structures, Exp Mech, 61 (2021), 431–443. https://doi.org/10.1007/s11340-020-00636-y doi: 10.1007/s11340-020-00636-y
|
| [27] |
S. Jin, Y. P. Korkolis, Y. Li, Shear resistance of an auxetic chiral mechanical metamaterial, Int J Solid Struct, 174 (2019), 28–37. https://doi.org/10.1016/j.ijsolstr.2019.06.005 doi: 10.1016/j.ijsolstr.2019.06.005
|
| [28] | N. Karathanasopoulos, F. Dos Reis, M. Diamantopoulou, J. F. Ganghoffer, Mechanics of beams made from chiral metamaterials: Tuning deflections through normal-shear strain couplings, Mater Des, 189 (2020), 108520. |
| [29] |
M. Khezri, K. Rasmussen, Functionalising buckling for structural morphing in kinetic façades: Concepts, strategies and applications, Thin Wall Struct, 180 (2022), 109749. https://doi.org/10.1016/j.matdes.2020.108520 doi: 10.1016/j.matdes.2020.108520
|
| [30] |
J. A. Kraus, J. J. Rimoli, An elastica theory for compressible imperfect beams with application to mechanical metamaterials, Mech Res Commun, 131 (2023), 104147. https://doi.org/10.1016/j.mechrescom.2023.104147 doi: 10.1016/j.mechrescom.2023.104147
|
| [31] |
R. Luciano, H. Darban, C. Bartolomeo, F. Fabbrocino, D. Scorza, Free flexural vibrations of nanobeams with non-classical boundary conditions using stress-driven nonlocal model, Mech Res Commun, 107 (2020), 103536. https://doi.org/10.1016/j.mechrescom.2020.103536 doi: 10.1016/j.mechrescom.2020.103536
|
| [32] |
G. Mancusi, F. Fabbrocino, L. Feo, F. Fraternali, Size effect and dynamic properties of 2D lattice materials, Compos Part B-eng, 112 (2017), 235–242. https://doi.org/10.1016/j.compositesb.2016.12.026 doi: 10.1016/j.compositesb.2016.12.026
|
| [33] |
P. K. Masjedi, P. M. Weaver, Analytical solution for arbitrary large deflection of geometrically exact beams using the homotopy analysis method, Appl Math Model, 103 (2022), 516–542. https://doi.org/10.1016/j.apm.2021.10.037 doi: 10.1016/j.apm.2021.10.037
|
| [34] |
A. Nuñez-Labielle, J. Cante, A. Huespe, J. Oliver, Towards shock absorbing hyperelastic metamaterial design. (i) macroscopic scale: Computational shock-capturing, Comput. Methods Appl. Mech. Eng., 393 (2022), 114732. https://doi.org/10.1016/j.cma.2022.114732 doi: 10.1016/j.cma.2022.114732
|
| [35] |
L. Paleari, M. Bragaglia, F. Fabbrocino, R. Luciano, F. Nanni, Self-monitoring performance of 3d-printed poly-ether-ether-ketone carbon nanotube composites, Polymers, 15 (2022), 8. https://doi.org/10.3390/polym15010008 doi: 10.3390/polym15010008
|
| [36] |
D. Qi, P. Zhang, W. Wu, K. Xin, H. Liao, Y. Li, et al., Innovative 3D chiral metamaterials under large deformation: Theoretical and experimental analysis, Int J Solids Struct, 202 (2020), 787–797. https://doi.org/10.1016/j.ijsolstr.2020.06.047 doi: 10.1016/j.ijsolstr.2020.06.047
|
| [37] |
A. Rafsanjani, K. Bertoldi, A. R. Studart, Programming soft robots with flexible mechanical metamaterials, Sci. Robot., 4 (2019), eaav7874. https://doi.org/10.1126/scirobotics.aav7874 doi: 10.1126/scirobotics.aav7874
|
| [38] |
H. Reda, S. Alavi, M. Nasimsobhan, J. Ganghoffer, Homogenization towards chiral cosserat continua and applications to enhanced timoshenko beam theories, Mech Mater, 155 (2021), 103728. https://doi.org/10.1016/j.mechmat.2020.103728 doi: 10.1016/j.mechmat.2020.103728
|
| [39] |
E. Riks, The Application of Newton's Method to the Problem of Elastic Stability, J. Appl. Mech., 39 (1972), 1060–1065. https://doi.org/10.1115/1.3422829 doi: 10.1115/1.3422829
|
| [40] |
P. Sengsri, S. Kaewunruen, Additive manufacturing meta-functional composites for engineered bridge bearings: A review, Constr Build Mater, 262 (2020), 120535. https://doi.org/10.1115/1.3422829 doi: 10.1115/1.3422829
|
| [41] |
A. Skrzat, V. A. Eremeyev, On the effective properties of foams in the framework of the couple stress theory, Continuum Mech. Thermodyn., 32 (2020), 1779–1801. https://doi.org/10.1007/s00161-020-00880-6 doi: 10.1007/s00161-020-00880-6
|
| [42] |
J. U. Surjadi, L. Gao, H. Du, X. Li, X. Xiong, N. X. Fang, et al., Mechanical metamaterials and their engineering applications, Adv Eng Mater, 21 (2019), 1800864. https://doi.org/10.1002/adem.201800864 doi: 10.1002/adem.201800864
|
| [43] |
E. Turco, E. Barchiesi, I. Giorgio, F. dell'Isola, A lagrangian hencky-type non-linear model suitable for metamaterials design of shearable and extensible slender deformable bodies alternative to timoshenko theory, Int. J. Non Linear Mech, 123 (2020), 103481. https://doi.org/10.1016/j.ijnonlinmec.2020.103481 doi: 10.1016/j.ijnonlinmec.2020.103481
|
| [44] |
E. N. Vilchevskaya, W. Müller, V. A. Eremeyev, Extended micropolar approach within the framework of 3M theories and variations thereof, Continuum Mech. Thermodyn., 34 (2022), 533–554. https://doi.org/10.1007/s00161-021-01072-6 doi: 10.1007/s00161-021-01072-6
|
| [45] | J. Wang, S. Zhu, L. Chen, T. Liu, H. Liu, Z. Lv, et al., Data mining from a hierarchical dataset for mechanical metamaterials composed of curved-sides triangles, Compos Struct, 319 (2023), 117153. |
| [46] |
O. Weeger, Numerical homogenization of second gradient, linear elastic constitutive models for cubic 3D beam-lattice metamaterials, Int J Solids Struct, 224 (2021), 111037. https://doi.org/10.1016/j.ijsolstr.2021.03.024 doi: 10.1016/j.ijsolstr.2021.03.024
|
| [47] |
W. Wu, P. Liu, Z. Kang, A novel mechanical metamaterial with simultaneous stretching- and compression-expanding property, Mater Des, 208 (2021), 109930. https://doi.org/10.1016/j.matdes.2021.109930 doi: 10.1016/j.matdes.2021.109930
|
| [48] |
B. Yang, M. Bacciocchi, N. Fantuzzi, R. Luciano, F. Fabbrocino, Wave propagation in periodic nano structures through second strain gradient elasticity, Int. J. Mech. Sci. Struct, 260 (2023), 108639. https://doi.org/10.1016/j.ijmecsci.2023.108639 doi: 10.1016/j.ijmecsci.2023.108639
|
| [49] |
B. Yang, M. Bacciocchi, N. Fantuzzi, R. Luciano, F. Fabbrocino, Computational simulation and acoustic analysis of two-dimensional nano-waveguides considering second strain gradient effects, Comput Struct, 296 (2024), 107299. https://doi.org/10.1016/j.compstruc.2024.107299 doi: 10.1016/j.compstruc.2024.107299
|
| [50] |
P. Zhang, D. Qi, R. Xue, K. Liu, W. Wu, Y. Li, Mechanical design and energy absorption performances of rational gradient lattice metamaterials, Compos Struct, 277 (2021), 114606. https://doi.org/10.1016/j.compstruct.2021.114606 doi: 10.1016/j.compstruct.2021.114606
|
| [51] |
X. Y. Zhang, X. Ren, Y. Zhang, Y. M. Xie, A novel auxetic metamaterial with enhanced mechanical properties and tunable auxeticity, Thin Wall Struct, 174 (2022), 109162. https://doi.org/10.1016/j.tws.2022.109162 doi: 10.1016/j.tws.2022.109162
|
| [52] |
S. Zolfaghari, D. Mostofinejad, N. Fantuzzi, R. Luciano, F. Fabbrocino, Experimental evaluation of FRP-concrete bond using externally-bonded reinforcement on grooves (EBROG) method, Compos Struct, 310 (2023), 116693. https://doi.org/10.1016/j.compstruct.2023.116693 doi: 10.1016/j.compstruct.2023.116693
|














Figures(15)
Jose Manuel Torres Espino, Emilio Barchiesi. Computational study of a homogenized nonlinear generalization of Timoshenko beam proposed by Turco et al.[J]. Networks and Heterogeneous Media, 2024, 19(3): 1133-1155. doi: 10.3934/nhm.2024050







 DownLoad:
DownLoad: