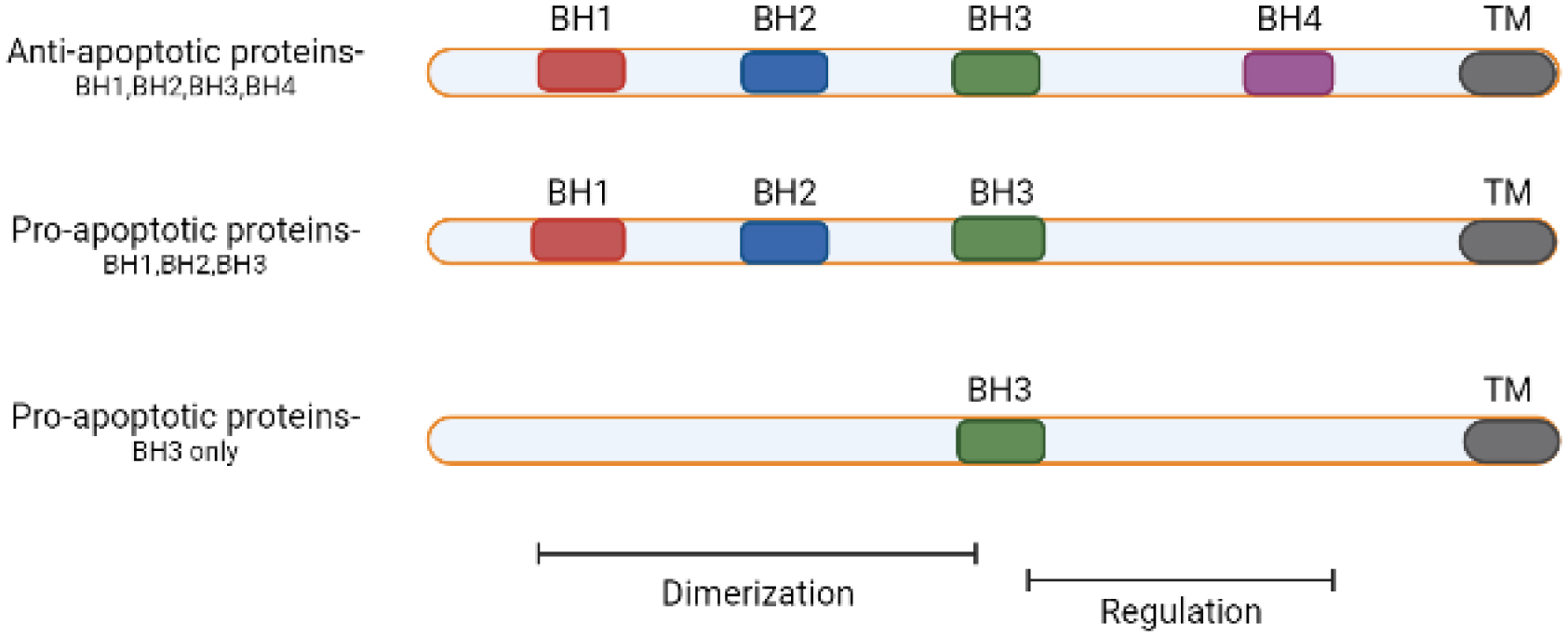
Breast cancer, a prominent form of cancer in women, arises from the inner lining of mammary glands, ducts, and lobules. With an approximate prevalence rate of 1 in 8 women, the standard treatment methods for this condition include the surgical excision of afflicted tissues, chemotherapy, radiation, and hormone therapy. The BCL-2 gene, also known as the B cell lymphoma gene, prevents apoptosis in eukaryotic cells. It is commonly found to be excessively active in many types of malignancies, such as leukemia, carcinomas, and breast cancer. The excessive expression of this gene has a role in the advancement of cancer by inhibiting apoptosis. Recent research emphasizes the function of microRNAs (miRs) in regulating the expression of BCL-2. These miRs can either decrease or increase the activity of specific genes involved in programmed cell death, thus making them potential targets for therapeutic interventions. This review explicitly examines the regulatory impacts of several miRs on BCL-2, thereby investigating their ability to trigger apoptosis and function as targeted treatments for breast cancer. By comprehending the complex interplay between miRs and BCL-2, it is possible to devise novel therapeutic approaches that can augment the efficacy of breast cancer treatments, thus eventually enhancing patient outcomes.
Citation: Kannan Mayuri, Sundaram Vickram, Thirunavukarasou Anand, Konda Mani Saravanan. MicroRNA-mediated regulation of BCL-2 in breast cancer[J]. AIMS Molecular Science, 2025, 12(1): 32-48. doi: 10.3934/molsci.2025003
Breast cancer, a prominent form of cancer in women, arises from the inner lining of mammary glands, ducts, and lobules. With an approximate prevalence rate of 1 in 8 women, the standard treatment methods for this condition include the surgical excision of afflicted tissues, chemotherapy, radiation, and hormone therapy. The BCL-2 gene, also known as the B cell lymphoma gene, prevents apoptosis in eukaryotic cells. It is commonly found to be excessively active in many types of malignancies, such as leukemia, carcinomas, and breast cancer. The excessive expression of this gene has a role in the advancement of cancer by inhibiting apoptosis. Recent research emphasizes the function of microRNAs (miRs) in regulating the expression of BCL-2. These miRs can either decrease or increase the activity of specific genes involved in programmed cell death, thus making them potential targets for therapeutic interventions. This review explicitly examines the regulatory impacts of several miRs on BCL-2, thereby investigating their ability to trigger apoptosis and function as targeted treatments for breast cancer. By comprehending the complex interplay between miRs and BCL-2, it is possible to devise novel therapeutic approaches that can augment the efficacy of breast cancer treatments, thus eventually enhancing patient outcomes.

| [1] |
Wilkinson L, Gathani T (2022) Understanding breast cancer as a global health concern. Br J Radiol 95: 20211033. https://doi.org/10.1259/bjr.20211033 
|
| [2] |
Bland KA, Mustafa R, McTaggart-Cowan H (2023) Patient preferences in metastatic breast cancer care: A scoping review. Cancers 15: 4331. https://doi.org/10.3390/cancers15174331
|
| [3] |
Bhushan A, Gonsalves A, Menon JU (2021) Current state of breast cancer diagnosis, treatment, and theranostics. Pharmaceutics 13: 723. https://doi.org/10.3390/pharmaceutics13050723
|
| [4] |
Pankotai-Bodó G, Oláh-Németh O, Sükösd F, et al. (2024) Routine molecular applications and recent advances in breast cancer diagnostics. J Biotechnol 380: 20-28. https://doi.org/10.1016/j.jbiotec.2023.12.005
|
| [5] |
Amethiya Y, Pipariya P, Patel S, et al. (2022) Comparative analysis of breast cancer detection using machine learning and biosensors. Intell Med 2: 69-81. https://doi.org/10.1016/j.imed.2021.08.004
|
| [6] |
Kandhavelu J, Subramanian K, Naidoo V, et al. (2024) A novel EGFR inhibitor, HNPMI, regulates apoptosis and oncogenesis by modulating BCL-2/BAX and p53 in colon cancer. Br J Pharmacol 181: 107-124. https://doi.org/10.1111/bph.16141
|
| [7] |
Sundaram KKM, Bupesh G, Saravanan KM (2022) Instrumentals behind embryo and cancer: A platform for prospective future in cancer research. AIMS Mol Sci 9: 25-45. https://doi.org/10.3934/molsci.2022002
|
| [8] |
Feng Y, Spezia M, Huang S, et al. (2018) Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis 5: 77-106. https://doi.org/10.1016/j.gendis.2018.05.001
|
| [9] |
Qian S, Wei Z, Yang W, et al. (2022) The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front Oncol 12: 985363. https://doi.org/10.3389/fonc.2022.985363
|
| [10] |
Debela DT, Muzazu SG, Heraro KD, et al. (2021) New approaches and procedures for cancer treatment: Current perspectives. SAGE Open Med 9: 20503121211034366. https://doi.org/10.1177/20503121211034366
|
| [11] |
Carlson RW, Anderson BO, Chopra R, et al. (2003) Treatment of breast cancer in countries with limited resources. Breast J 9: S67-S74. https://doi.org/10.1046/j.1524-4741.9.s2.6.x
|
| [12] |
Bamodu OA, Chung CC, Pisanic TR, et al. (2024) The intricate interplay between cancer stem cells and cell-of-origin of cancer: Implications for therapeutic strategies. Front Oncol 14: 1404628. https://doi.org/10.3389/fonc.2024.1404628
|
| [13] |
Bhat GR, Sethi I, Sadida HQ, et al. (2024) Cancer cell plasticity: From cellular, molecular, and genetic mechanisms to tumor heterogeneity and drug resistance. Cancer Metastasis Rev 43: 197-228. https://doi.org/10.1007/s10555-024-10172-z
|
| [14] |
Galluzzi L, Vitale I, Aaronson SA, et al. (2018) Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ 25: 486-541. https://doi.org/10.1038/s41418-017-0012-4
|
| [15] |
Warren CFA, Wong-Brown MW, Bowden NA (2019) BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis 10: 177. https://doi.org/10.1038/s41419-019-1407-6
|
| [16] |
Rabkin CS, Hirt C, Janz S, et al. (2008) t(14;18) Translocations and risk of follicular lymphoma. JNCI Monogr 2008: 48-51. https://doi.org/10.1093/jncimonographs/lgn002
|
| [17] |
Roulland S, Lebailly P, Lecluse Y, et al. (2004) Characterization of the t(14;18) BCL2-IGH translocation in farmers occupationally exposed to pesticides. Cancer Res 64: 2264-2269. https://doi.org/10.1158/0008-5472.can-03-3604
|
| [18] |
Singh R, Letai A, Sarosiek K (2019) Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol 20: 175-193. https://doi.org/10.1038/s41580-018-0089-8
|
| [19] |
Banjara S, Suraweera CD, Hinds MG, et al. (2020) The BCL-2 family: Ancient origins, conserved structures, and divergent mechanisms. Biomolecules 10: 128. https://doi.org/10.3390/biom10010128
|
| [20] |
Zografos E, Zagouri F, Kalapanida D, et al. (2019) Prognostic role of microRNAs in breast cancer: A systematic review. Oncotarget 10: 7156-7178. https://doi.org/10.18632/oncotarget.27327
|
| [21] |
Muñoz JP, Pérez-Moreno P, Pérez Y, et al. (2023) The role of microRNAs in breast cancer and the challenges of their clinical application. Diagnostics 13: 3072. https://doi.org/10.3390/diagnostics13193072
|
| [22] |
Sharma PC, Gupta A (2020) MicroRNAs: Potential biomarkers for diagnosis and prognosis of different cancers. Transl Cancer Res 9: 5798-5818. https://doi.org/10.21037/tcr-20-1294
|
| [23] |
Ying SY, Chang DC, Lin SL (2008) The microRNA (miRNA): Overview of the RNA genes that modulate gene function. Mol Biotechnol 38: 257-268. https://doi.org/10.1007/s12033-007-9013-8
|
| [24] |
Pekarek L, Torres-Carranza D, Fraile-Martinez O, et al. (2023) An overview of the role of microRNAs on carcinogenesis: A focus on cell cycle, angiogenesis and metastasis. Int J Mol Sci 24: 7268. https://doi.org/10.3390/ijms24087268
|
| [25] |
Sharifi Z, Talkhabi M, Taleahmad S (2022) Identification of potential microRNA diagnostic panels and uncovering regulatory mechanisms in breast cancer pathogenesis. Sci Rep 12: 20135. https://doi.org/10.1038/s41598-022-24347-7
|
| [26] |
Abdul Manap AS, Wisham AA, Wong FW, et al. (2024) Mapping the function of microRNAs as a critical regulator of tumor-immune cell communication in breast cancer and potential treatment strategies. Front Cell Dev Biol 12: 1390704. https://doi.org/10.3389/fcell.2024.1390704
|
| [27] |
Kaloni D, Diepstraten ST, Strasser A, et al. (2023) BCL-2 protein family: Attractive targets for cancer therapy. Apoptosis 28: 20-38. https://doi.org/10.1007/s10495-022-01780-7
|
| [28] |
O'Day E, Lal A (2010) MicroRNAs and their target gene networks in breast cancer. Breast Cancer Res 12: 201. https://doi.org/10.1186/bcr2484
|
| [29] |
Saddam M, Paul SK, Habib MA, et al. (2024) Emerging biomarkers and potential therapeutics of the BCL-2 protein family: The apoptotic and antiapoptotic context. Egypt J Med Hum Genet 25: 12. https://doi.org/10.1186/s43042-024-00485-7
|
| [30] |
Wang H, Guo M, Wei H, et al. (2023) Targeting p53 pathways: Mechanisms, structures, and advances in therapy. Sig Transduct Target Ther 8: 92. https://doi.org/10.1038/s41392-023-01347-1
|
| [31] |
Elmore S (2007) Apoptosis: A review of programmed cell death. Toxicol Pathol 35: 495-516. https://doi.org/10.1080/01926230701320337
|
| [32] |
Jan R, Chaudhry G (2019) Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv Pharm Bull 9: 205-218. https://doi.org/10.15171/apb.2019.024
|
| [33] |
Kale J, Osterlund EJ, Andrews DW (2018) BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ 25: 65-80. https://doi.org/10.1038/cdd.2017.186
|
| [34] |
Letai A, Bassik MC, Walensky LD, et al. (2002) Distinct BH3 domains either sensitize or activate mitochondrial apoptosis, serving as prototype cancer therapeutics. Cancer Cell 2: 183-192. https://doi.org/10.1016/S1535-6108(02)00127-7
|
| [35] |
Lomonosova E, Chinnadurai G (2008) BH3-only proteins in apoptosis and beyond: An overview. Oncogene 27: S2-S19. https://doi.org/10.1038/onc.2009.39
|
| [36] |
Lalier L, Vallette F, Manon S (2022) BCL-2 family members and the mitochondrial import machineries: The roads to death. Biomolecules 12: 162. https://doi.org/10.3390/biom12020162
|
| [37] |
Chen D, Frezza M, Schmitt S, et al. (2011) Bortezomib as the first proteasome inhibitor anticancer drug: Current status and future perspectives. Curr Cancer Drug Targets 11: 239-253. https://doi.org/10.2174/156800911794519752
|
| [38] |
Harris MH, Thompson CB (2000) The role of the BCL-2 family in the regulation of outer mitochondrial membrane permeability. Cell Death Differ 7: 1182-1191. https://doi.org/10.1038/sj.cdd.4400781
|
| [39] |
Jang DM, Oh EK, Hahn H, et al. (2022) Structural insights into apoptotic regulation of human Bfk as a novel BCL-2 family member. Comput Struct Biotechnol J 20: 745-756. https://doi.org/10.1016/j.csbj.2022.01.023
|
| [40] |
Kelekar A, Thompson CB (1998) BCL-2-family proteins: The role of the BH3 domain in apoptosis. Trends Cell Biol 8: 324-330. https://doi.org/10.1016/s0962-8924(98)01321-x
|
| [41] |
Dadsena S, Jenner A, García-Sáez AJ (2021) Mitochondrial outer membrane permeabilization at the single molecule level. Cell Mol Life Sci 78: 3777-3790. https://doi.org/10.1007/s00018-021-03771-4
|
| [42] |
Bender T, Martinou JC (2013) Where killers meet—permeabilization of the outer mitochondrial membrane during apoptosis. Cold Spring Harb Perspect Biol 5: a011106. https://doi.org/10.1101/cshperspect.a011106
|
| [43] |
Czabotar PE, Garcia-Saez AJ (2023) Mechanisms of BCL-2 family proteins in mitochondrial apoptosis. Nat Rev Mol Cell Biol 24: 732-748. https://doi.org/10.1038/s41580-023-00629-4
|
| [44] |
Peña-Blanco A, García-Sáez AJ (2018) Bax, Bak and beyond—mitochondrial performance in apoptosis. FEBS J 285: 416-431. https://doi.org/10.1111/febs.14186
|
| [45] |
MacFarlane LA, Murphy PR (2010) MicroRNA: Biogenesis, function and role in cancer. Curr Genomics 11: 537-561. https://doi.org/10.2174/138920210793175895
|
| [46] |
Starega-Roslan J, Koscianska E, Kozlowski P, et al. (2011) The role of the precursor structure in the biogenesis of microRNA. Cell Mol Life Sci 68: 2859-2871. https://doi.org/10.1007/s00018-011-0726-2
|
| [47] |
O'Brien J, Hayder H, Zayed Y, et al. (2018) Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol 9: 402. https://doi.org/10.3389/fendo.2018.00402
|
| [48] |
Haas G, Cetin S, Messmer M, et al. (2016) Identification of factors involved in target RNA-directed microRNA degradation. Nucleic Acids Res 44: 2873-2887. https://doi.org/10.1093/nar/gkw040
|
| [49] | Bujak M, Ratkaj I, Baus M, et al. (2011) Small interfering RNAs: Heralding a new era in gene therapy. Gene therapy—developments and future perspectives . IntechOpen. https://doi.org/10.5772/20646 |
| [50] |
Gu S, Jin L, Zhang F, et al. (2009) Biological basis for restriction of microRNA targets to the 3′ untranslated region in mammalian mRNAs. Nat Struct Mol Biol 16: 144-150. https://doi.org/10.1038/nsmb.1552
|
| [51] |
Menon A, Abd-Aziz N, Khalid K, et al. (2022) miRNA: A promising therapeutic target in cancer. Int J Mol Sci 23: 11502. https://doi.org/10.3390/ijms231911502
|
| [52] |
Peng Y, Croce CM (2016) The role of microRNAs in human cancer. Signal Transduct Target Ther 1: 15004. https://doi.org/10.1038/sigtrans.2015.4
|
| [53] |
Otmani K, Lewalle P (2021) Tumor suppressor miRNA in cancer cells and the tumor microenvironment: Mechanism of deregulation and clinical implications. Front Oncol 11: 708765. https://doi.org/10.3389/fonc.2021.708765
|
| [54] |
Spadaccino F, Gigante M, Netti GS, et al. (2021) The ambivalent role of miRNAs in carcinogenesis: Involvement in renal cell carcinoma and their clinical applications. Pharmaceuticals 14: 322. https://doi.org/10.3390/ph14040322
|
| [55] |
Shah MY, Ferrajoli A, Sood AK, et al. (2016) microRNA therapeutics in cancer—An emerging concept. EBioMedicine 12: 34-42. https://doi.org/10.1016/j.ebiom.2016.09.017
|
| [56] |
Bayir H, Kagan VE (2008) Bench-to-bedside review: Mitochondrial injury, oxidative stress and apoptosis—there is nothing more practical than a good theory. Crit Care 12: 206. https://doi.org/10.1186/cc6779
|
| [57] |
Garrido C, Galluzzi L, Brunet M, et al. (2006) Mechanisms of cytochrome c release from mitochondria. Cell Death Differ 13: 1423-1433. https://doi.org/10.1038/sj.cdd.4401950
|
| [58] |
Adams JM, Cory S (2018) The BCL-2 arbiters of apoptosis and their growing role as cancer targets. Cell Death Differ 25: 27-36. https://doi.org/10.1038/cdd.2017.161
|
| [59] |
Garrido-Cano I, Pattanayak B, Adam-Artigues A, et al. (2022) MicroRNAs as a clue to overcome breast cancer treatment resistance. Cancer Metastasis Rev 41: 77-105. https://doi.org/10.1007/s10555-021-09992-0
|
| [60] |
Chakrabortty A, Patton DJ, Smith BF, et al. (2023) miRNAs: Potential as biomarkers and therapeutic targets for cancer. Genes 14: 1375. https://doi.org/10.3390/genes14071375
|
| [61] |
Hong T, Ding J, Li W (2019) mir-7 reverses breast cancer resistance to chemotherapy by targeting MRP1 and BCL2. Onco Targets Ther 12: 11097-11105. https://doi.org/10.2147/OTT.S213780
|
| [62] |
Cittelly DM, Das PM, Salvo VA, et al. (2010) Oncogenic HER2Δ16 suppresses miR-15a/16 and deregulates BCL-2 to promote endocrine resistance of breast tumors. Carcinogenesis 31: 2049-2057. https://doi.org/10.1093/carcin/bgq192
|
| [63] |
Tao L, Wu YQ, Zhang SP (2019) MiR-21-5p enhances the progression and paclitaxel resistance in drug-resistant breast cancer cell lines by targeting PDCD4. Neoplasma 66: 746-755. https://doi.org/10.4149/neo_2018_181207N930
|
| [64] |
Manvati S, Mangalhara KC, Kalaiarasan P, et al. (2015) miR-24-2 regulates genes in survival pathway and demonstrates potential in reducing cellular viability in combination with docetaxel. Gene 567: 217-224. https://doi.org/10.1016/j.gene.2015.05.003
|
| [65] |
Han X, Li Q, Liu C, et al. (2019) Overexpression miR-24-3p repressed Bim expression to confer tamoxifen resistance in breast cancer. J Cell Biochem 120: 12966-12976. https://doi.org/10.1002/jcb.28568
|
| [66] |
Körner C, Keklikoglou I, Bender C, et al. (2013) MicroRNA-31 sensitizes human breast cells to apoptosis by direct targeting of protein Kinase C ϵ (PKCϵ). J Biol Chem 288: 8750-8761.
|
| [67] |
Kastl L, Brown I, Schofield AC (2012) miRNA-34a is associated with docetaxel resistance in human breast cancer cells. Breast Cancer Res Treat 131: 445-454. https://doi.org/10.1007/s10549-011-1424-3
|
| [68] |
Bao C, Chen J, Chen D, et al. (2020) MiR-93 suppresses tumorigenesis and enhances chemosensitivity of breast cancer via dual targeting E2F1 and CCND1. Cell Death Dis 11: 618. https://doi.org/10.1038/s41419-020-02855-6
|
| [69] |
You F, Luan H, Sun D, et al. (2018) miRNA-106a promotes breast cancer cell proliferation, clonogenicity, migration, and invasion through inhibiting apoptosis and chemosensitivity. DNA Cell Biol 38: 198-207. https://doi.org/10.1089/dna.2018.4282
|
| [70] |
Zhang W, Jiang H, Chen Y, et al. (2019) Resveratrol chemosensitizes adriamycin-resistant breast cancer cells by modulating miR-122-5p. J Cell Biochem 120: 16283-16292. https://doi.org/10.1002/jcb.28910
|
| [71] |
Xie X, Hu Y, Xu L, et al. (2015) The role of miR-125b-mitochondria-caspase-3 pathway in doxorubicin resistance and therapy in human breast cancer. Tumor Biol 36: 7185-7194. https://doi.org/10.1007/s13277-015-3438-7
|
| [72] |
Zhou M, Liu Z, Zhao Y, et al. (2010) MicroRNA-125b confers the resistance of breast cancer cells to paclitaxel through suppression of pro-apoptotic BCL-2 antagonist killer 1 (Bak1) expression. J Biol Chem 285: 21496-21507. https://doi.org/10.1074/jbc.M109.083337
|
| [73] |
O'Brien K, Lowry MC, Corcoran C, et al. (2015) miR-134 in extracellular vesicles reduces triple-negative breast cancer aggression and increases drug sensitivity. Oncotarget 6: 32774-32789. https://doi.org/10.18632/oncotarget.5192
|
| [74] |
Xiang F, Fan Y, Ni Z, et al. (2019) Ursolic acid reverses the chemoresistance of breast cancer cells to paclitaxel by targeting miRNA-149-5p/MyD88. Front Oncol 9: 00501. https://doi.org/10.3389/fonc.2019.00501
|
| [75] |
Zheng Y, Lv X, Wang X, et al. (2016) miR-181b promotes chemoresistance in breast cancer by regulating Bim expression. Oncol Rep 35: 683-690. https://doi.org/10.3892/or.2015.4417
|
| [76] |
Zhang Y, He Y, Lu LL, et al. (2019) miRNA-192-5p impacts the sensitivity of breast cancer cells to doxorubicin via targeting peptidylprolyl isomerase A. Kaohsiung J Med Sci 35: 17-23. https://doi.org/10.1002/kjm2.12004
|
| [77] |
Yang G, Wu D, Zhu J, et al. (2013) Upregulation of miR-195 increases the sensitivity of breast cancer cells to Adriamycin treatment through inhibition of Raf-1. Oncol Rep 30: 877-889. https://doi.org/10.3892/or.2013.2532
|
| [78] |
Aakko S, Straume AH, Birkeland EE, et al. (2019) MYC-induced miR-203b-3p and miR-203a-3p control BCL-xL expression and paclitaxel sensitivity in tumor cells. Transl Oncol 12: 170-179. https://doi.org/10.1016/j.tranon.2018.10.001
|
| [79] |
Ye Z, Hao R, Cai Y, et al. (2016) Knockdown of miR-221 promotes the cisplatin-inducing apoptosis by targeting the BIM-Bax/Bak axis in breast cancer. Tumor Biol 37: 4509-4515. https://doi.org/10.1007/s13277-015-4267-4
|
| [80] |
Dai H, Xu L, Qian Q, et al. (2019) MicroRNA-222 promotes drug resistance to doxorubicin in breast cancer via regulation of miR-222/bim pathway. Biosci Rep 39: BSR20190650. https://doi.org/10.1042/BSR20190650
|
| [81] |
Gu X, Li JY, Guo J, et al. (2015) Influence of MiR-451 on drug resistances of paclitaxel-resistant breast cancer cell line. Med Sci Monit 21: 3291-3297. https://doi.org/10.12659/msm.894475
|
| [82] |
Chen X, Wang YW, Xing AY, et al. (2016) Suppression of SPIN1-mediated PI3K–Akt pathway by miR-489 increases chemosensitivity in breast cancer. J Pathol 239: 459-472. https://doi.org/10.1002/path.4743
|
| [83] | Jiang H, Cheng L, Hu P, et al. (2018) MicroRNA‑663b mediates TAM resistance in breast cancer by modulating TP73 expression. Mol Med Rep 18: 1120-1126. https://doi.org/10.3892/mmr.2018.9064 |
| [84] |
Ito Y, Inoue A, Seers T, et al. (2017) Identification of targets of tumor suppressor microRNA-34a using a reporter library system. Proc Natl Acad Sci 114: 3927-3932. https://doi.org/10.1073/pnas.1620019114
|
| [85] |
Cimmino A, Calin GA, Fabbri M, et al. (2005) miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci U S A 102: 13944-13949. https://doi.org/10.1073/pnas.0506654102
|
| [86] |
Sims EK, Lakhter AJ, Anderson-Baucum E, et al. (2017) MicroRNA 21 targets BCL2 mRNA to increase apoptosis in rat and human beta cells. Diabetologia 60: 1057-1065. https://doi.org/10.1007/s00125-017-4237-z
|
| [87] |
Garrido-Palacios A, Rojas Carvajal AM, Núñez-Negrillo AM, et al. (2023) MicroRNA dysregulation in early breast cancer diagnosis: A systematic review and meta-analysis. Int J Mol Sci 24: 8270. https://doi.org/10.3390/ijms24098270
|
| [88] |
Capaccia C, Diverio S, Zampini D, et al. (2022) The complex interaction between P53 and miRNAs joins new awareness in physiological stress responses. Cells 11: 1631. https://doi.org/10.3390/cells11101631
|
| [89] |
Arif KMT, Elliott EK, Haupt LM, et al. (2020) Regulatory mechanisms of epigenetic miRNA relationships in human cancer and potential as therapeutic targets. Cancers 12: 2922. https://doi.org/10.3390/cancers12102922
|
| [90] |
Calin GA, Dumitru CD, Shimizu M, et al. (2002) Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A 99: 15524-15529. https://doi.org/10.1073/pnas.242606799
|
| [91] |
Peters L, Meister G (2007) Argonaute proteins: Mediators of RNA silencing. Mol Cell 26: 611-623. https://doi.org/10.1016/j.molcel.2007.05.001
|
| [92] |
Ma B, Wang S, Wu W, et al. (2023) Mechanisms of circRNA/lncRNA-miRNA interactions and applications in disease and drug research. Biomed Pharmacother 162: 114672. https://doi.org/10.1016/j.biopha.2023.114672
|
| [93] |
Fadeel B, Orrenius S (2005) Apoptosis: A basic biological phenomenon with wide-ranging implications in human disease. J Intern Med 258: 479-517. https://doi.org/10.1111/j.1365-2796.2005.01570.x
|
| [94] |
Si W, Shen J, Zheng H, et al. (2019) The role and mechanisms of action of microRNAs in cancer drug resistance. Clin Epigenet 11: 25. https://doi.org/10.1186/s13148-018-0587-8
|
| [95] |
Mohammad RM, Muqbil I, Lowe L, et al. (2015) Broad targeting of resistance to apoptosis in cancer. Semin Cancer Biol 35: S78-S103. https://doi.org/10.1016/j.semcancer.2015.03.001
|
| [96] | Callens M, Kraskovskaya N, Derevtsova K, et al. (2021) The role of BCL-2 proteins in modulating neuronal Ca2+ signaling in health and in Alzheimer's disease. BBA-Mol Cell Res 1868: 118997. https://doi.org/10.1016/j.bbamcr.2021.118997 |
| [97] |
D'Aguanno S, Brignone M, Scalera S, et al. (2024) BCL-2 dependent modulation of Hippo pathway in cancer cells. Cell Commun Signal 22: 277. https://doi.org/10.1186/s12964-024-01647-1
|
| [98] | Abaza A, Vasavada AM, Sadhu A, et al. (2022) A systematic review of apoptosis in correlation with cancer: Should apoptosis be the ultimate target for cancer treatment?. Cureus 14: e28496. https://doi.org/10.7759/cureus.28496 |
| [99] | Huska J, Lamb HM, Hardwick JM (2019) Overview of BCL-2 family proteins and therapeutic potentials. BCL-2 family proteins. Methods in molecular biology . New York: Humana Press 1-21. https://doi.org/10.1007/978-1-4939-8861-7_1 |
| [100] |
Yap JL, Chen L, Lanning ME, et al. (2017) Expanding the cancer arsenal with targeted therapies: Disarmament of the antiapoptotic BCL-2 proteins by small molecules. J Med Chem 60: 821-838. https://doi.org/10.1021/acs.jmedchem.5b01888
|
| [101] |
Tassone P, Di Martino MT, Arbitrio M, et al. (2023) Safety and activity of the first-in-class locked nucleic acid (LNA) miR-221 selective inhibitor in refractory advanced cancer patients: a first-in-human, phase 1, open-label, dose-escalation study. J Hematol Oncol 16: 68. https://doi.org/10.1186/s13045-023-01468-8
|
| [102] |
Qassem S, Breier D, Naidu GS, et al. (2024) Unlocking the therapeutic potential of locked nucleic acids through lipid nanoparticle delivery. Mol Ther Nucl Acids 35: 102224. https://doi.org/10.1016/j.omtn.2024.102224
|
| [103] |
Riolo G, Cantara S, Marzocchi C, et al. (2020) miRNA targets: From prediction tools to experimental validation. Methods Protoc 4: 1. https://doi.org/10.3390/mps4010001
|
| [104] |
Hersh AM, Alomari S, Tyler BM (2022) Crossing the blood-brain barrier: Advances in nanoparticle technology for drug delivery in neuro-oncology. Int J Mol Sci 23: 4153. https://doi.org/10.3390/ijms23084153
|


Figures(3) / Tables(1)

Kannan Mayuri, Sundaram Vickram, Thirunavukarasou Anand, Konda Mani Saravanan. MicroRNA-mediated regulation of BCL-2 in breast cancer[J]. AIMS Molecular Science, 2025, 12(1): 32-48. doi: 10.3934/molsci.2025003







 DownLoad:
DownLoad: