Citation: Jin Hwan Do. Apomorphine-induced pathway perturbation in MPP+-treated SH-SY5Y cells[J]. AIMS Molecular Science, 2017, 4(3): 271-287. doi: 10.3934/molsci.2017.3.271

| [1] |
Ribarič S (2012) The pharmacological properties and therapeutic use of apomorphine. Molecules 17: 5289-5309. |
| [2] |
Deleu D, Hanssens Y, Northway MG (2004) Subcutaneous apomorphine: an evidence-based review of its use in Parkinson's disease. Drugs Aging 21: 687-709. doi: 10.2165/00002512-200421110-00001

|
| [3] |
Sydow O (2008) Parkinson's disease: recent development in therapies for advanced disease with a focus on deep brain stimulation (DBS) and duodenal levodopa infusion. FEBS J 275: 1370-1376. doi: 10.1111/j.1742-4658.2008.06295.x
|
| [4] |
Haeri M, Sarbaz Y, Gharibzadeh S (2005) Modeling the Parkinson's tremor and its treatments. J Theor Biol 236: 311-322. doi: 10.1016/j.jtbi.2005.03.014
|
| [5] |
Antonini A, Isaias IU, Rodolfi G, et al. (2011) A 5-year prospective assessment of advanced Parkinson disease patients treated with subcutaneous apomorphine infusion or deep brain stimulation. J Neurol 258: 579-585. doi: 10.1007/s00415-010-5793-z
|
| [6] |
Carron R, Fraix V, Maineri C, et al. (2011) High frequency deep brain stimulation of the subthalamic nucleus versus continuous subcutaneous apomorphine infusion therapy: A review. J Neural Transm 118: 915-924. doi: 10.1007/s00702-010-0556-7
|
| [7] |
De Gaspari D, Siri C, Landi A, et al. (2006) Clinical and neuropsychological follow up at 12 months in patients with complicated Parkinson's disease treated with subcutaneous apomorphine infusion or deep brain stimulation of the subthalamic nucleus. J Neurol Neurosurg Psychiatry 77: 450-453. doi: 10.1136/jnnp.2005.078659
|
| [8] |
Merello M, Lees AJ, Balej J, et al. (1999) GPi firing rate modification during beginning-of-dose motor deterioration following acute administration of apomorphine. Mov Disord 14: 481-483. doi: 10.1002/1531-8257(199905)14:3<481::AID-MDS1015>3.0.CO;2-H
|
| [9] |
Gassen M, Glinka Y, Pinchasi B, et al. (1996) Apomorphine is a highly potent free radical scavenger in rat brain mitochondrial fraction. Eur J Pharmacol 308: 219-225. doi: 10.1016/0014-2999(96)00291-9
|
| [10] | Grunblatt E, Mandel S, Gassen M, et al. (1999) Potent neuroprotective and antioxidant activity of apomorphine in MPTP and 6-hydroxydopamine induced neurotoxicity. J Neural Transm Suppl 55: 57-70. |
| [11] |
Do JH (2014) Neurotoxin-induced pathway perturbation in human neuroblastoma SH-EP cells. Mol Cells 37: 672-684. doi: 10.14348/molcells.2014.0173
|
| [12] |
Kim IS, Choi DK, Do JH (2013) Genome-wide temporal responses of human neuroblastoma SH-SY5Y cells to MPP+ neurotoxity. BioChip J 7: 247-257. doi: 10.1007/s13206-013-7308-3
|
| [13] | Nakamura K, Bindokas VP, Marks JD, et al. (2000) The selective toxicity of 1-methyl-4-phenylpyridinium to dopaminergic neurons: The role of mitochondria complex I and reactive oxygen species revisited. Mol Pharmacol 58: 271-278. |
| [14] | Lotharius J, O'Malley KL (2000) The parkinsonisminducing drug 1-methyl-4-phenylpyridinium triggers intracellular dopamine oxidation. A novel mechanism of toxicity. J Biol Chem 275: 38581-38588. |
| [15] |
Moon HE, Paek SH (2015) Mitochondrial dysfunction in Parkinson's disease. Exp Neurobiol 24: 103-116. doi: 10.5607/en.2015.24.2.103
|
| [16] |
Choi DK, Kim IS, Do JH (2014) Signaling pathway analysis of MPP+-treated human neuroblastoma SH-SY5Y cells. Biotechnol Bioprocess Eng 19: 332-340. doi: 10.1007/s12257-013-0754-x
|
| [17] |
Du P, Kibbe WA, Lin SM (2008) Lumi: a pipeline for processing Illumina microarray. Bioinformatics 24: 1547-1548. doi: 10.1093/bioinformatics/btn224
|
| [18] |
Conesa A, Nueda MN, Ferrer A, et al. (2006) maSigPro: a method to identify significantly differential expression profiles in time-course microarray experiments. Bioinformatics 22: 1096-1102. doi: 10.1093/bioinformatics/btl056
|
| [19] |
Yu G, Wang LG, Han Y, et al. (2012) clusterProfiler: an R package for comparing biological themes among clusters. Omics 16: 284-287. doi: 10.1089/omi.2011.0118
|
| [20] |
Tarca AL, Draghici S, Khatri P, et al. (2009) A novel signaling pathway impact analysis. Bioinformatics 25: 75-82. doi: 10.1093/bioinformatics/btn577
|
| [21] | Csárdi G, Nepusz T (2006) The igraph software package for complex network research. Inter J Complex Syst 1695. |
| [22] |
Pepe D, Do JH (2015) Estimation of dysregulated pathway regions in MPP+ treated human neuroblastoma SH-EP cells with structural equation model. BioChip J 9: 131-138. doi: 10.1007/s13206-015-9206-3
|
| [23] | Pepe D, Do JH (2016) Comparison of perturbed pathways in two different cell models for Parkinson's disease with structural equation model. J Comput Biol 23: 90-101. |
| [24] |
Newman ME, Girvan M (2004) Finding and evaluating community structure in networks. Phys Rev E Stat Nonlin Soft Matter Phys 69: 026113. doi: 10.1103/PhysRevE.69.026113
|
| [25] |
Benjamini Y, Yekutieli D (2001) The control of the false discovery rate in multiple testing under dependency. Ann Stat 29: 1165-1188. doi: 10.1214/aos/1013699998
|
| [26] |
Yu G, Wang KG, Yan GR, et al. (2015) DOSE: an R/Bioconductor package for disease ontology semantic and enrichment analysis. Bioinformatics 31: 608-609. doi: 10.1093/bioinformatics/btu684
|
| [27] | Okun MS, Foote KD (2010) Parkinson's disease DBS: what, when, who and why? The time has come to tailor DBS targets. Expert Rev Neurother 10: 1847-1857. |
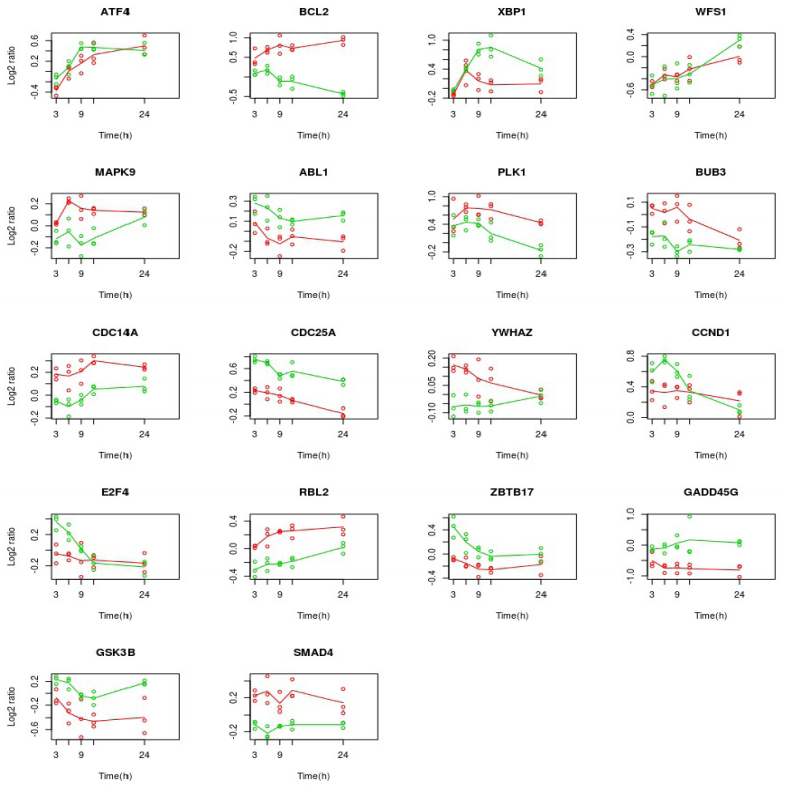
Figures(5) / Tables(4)

Jin Hwan Do. Apomorphine-induced pathway perturbation in MPP+-treated SH-SY5Y cells[J]. AIMS Molecular Science, 2017, 4(3): 271-287. doi: 10.3934/molsci.2017.3.271







 DownLoad:
DownLoad: