Citation: Kristyn Alissa Bates. Gene-environment interactions in considering physical activity for the prevention of dementia[J]. AIMS Molecular Science, 2015, 2(3): 359-381. doi: 10.3934/molsci.2015.3.359

| [1] |
Alzheimer A, Stelzmann RA, Schnitzlein HN, et al. (1995) An English translation of Alzheimer's 1907 paper, "Uber eine eigenartige Erkankung der Hirnrinde". Clin Anat 8: 429-431. doi: 10.1002/ca.980080612

|
| [2] |
(2013) 2013 Alzheimer's disease facts and figures. Alzheimers Dement 9: 208-245. doi: 10.1016/j.jalz.2013.02.003
|
| [3] |
Brookmeyer R, Johnson E, Ziegler-Graham K, et al. (2007) Forecasting the global burden of Alzheimer's disease. Alzheimers Dement 3: 186-191. doi: 10.1016/j.jalz.2007.04.381
|
| [4] |
Martins RN, Robinson PJ, Chleboun JO, et al. (1991) The molecular pathology of amyloid deposition in Alzheimer's disease. Mol Neurobiol 5: 389-398. doi: 10.1007/BF02935560
|
| [5] | Masters CL, Multhaup G, Simms G, et al. (1985) Neuronal origin of a cerebral amyloid: neurofibrillary tangles of Alzheimer's disease contain the same protein as the amyloid of plaque cores and blood vessels. Embo J 4: 2757-2763. |
| [6] |
Schweers O, Mandelkow EM, Biernat J, et al. (1995) Oxidation of cysteine-322 in the repeat domain of microtubule-associated protein tau controls the in vitro assembly of paired helical filaments. Proc Natl Acad Sci U S A 92: 8463-8467. doi: 10.1073/pnas.92.18.8463
|
| [7] |
Goedert M, Jakes R, Spillantini MG, et al. (1996) Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature 383: 550-553. doi: 10.1038/383550a0
|
| [8] |
Villemagne VL, Pike KE, Darby D, et al. (2008) Abeta deposits in older non-demented individuals with cognitive decline are indicative of preclinical Alzheimer's disease. Neuropsychologia 46: 1688-1697. doi: 10.1016/j.neuropsychologia.2008.02.008
|
| [9] | Chien DT, Bahri S, Szardenings AK, et al. (2013) Early clinical PET imaging results with the novel PHF-tau radioligand [F-18]-T807. J Alzheimers Dis 34: 457-468. |
| [10] |
Maruyama M, Shimada H, Suhara T, et al. (2013) Imaging of tau pathology in a tauopathy mouse model and in Alzheimer patients compared to normal controls. Neuron 79: 1094-1108. doi: 10.1016/j.neuron.2013.07.037
|
| [11] |
Rentz DM, Parra Rodriguez MA, Amariglio R, et al. (2013) Promising developments in neuropsychological approaches for the detection of preclinical Alzheimer's disease: a selective review. Alzheimers Res Ther 5: 58. doi: 10.1186/alzrt222
|
| [12] | Baumgart M, Snyder HM, Carrillo MC, et al. (2015) Summary of the evidence on modifiable risk factors for cognitive decline and dementia: A population-based perspective. Alzheimers Dement. |
| [13] |
Barnes DE, Yaffe K (2011) The projected effect of risk factor reduction on Alzheimer's disease prevalence. Lancet Neurol 10: 819-828. doi: 10.1016/S1474-4422(11)70072-2
|
| [14] |
Selkoe DJ (1996) Amyloid beta-protein and the genetics of Alzheimer's disease. J Biol Chem 271: 18295-18298. doi: 10.1074/jbc.271.31.18295
|
| [15] |
Glenner GG, Wong CW (1984) Alzheimer's disease and Down's syndrome: sharing of a unique cerebrovascular amyloid fibril protein. Biochem Biophys Res Commun 122: 1131-1135. doi: 10.1016/0006-291X(84)91209-9
|
| [16] |
Mann DM, Esiri MM (1989) The pattern of acquisition of plaques and tangles in the brains of patients under 50 years of age with Down's syndrome. J Neurol Sci 89: 169-179. doi: 10.1016/0022-510X(89)90019-1
|
| [17] |
Jorm AF, Korten AE, Henderson AS (1987) The prevalence of dementia: a quantitative integration of the literature. Acta Psychiatr Scand 76: 465-479. doi: 10.1111/j.1600-0447.1987.tb02906.x
|
| [18] |
Hebert LE, Weuve J, Scherr PA, et al. (2013) Alzheimer disease in the United States (2010-2050) estimated using the 2010 census. Neurology 80: 1778-1783. doi: 10.1212/WNL.0b013e31828726f5
|
| [19] |
(2015) 2015 Alzheimer's disease facts and figures. Alzheimers Dement 11: 332-384. doi: 10.1016/j.jalz.2015.02.003
|
| [20] |
Lukiw WJ (2004) Gene expression profiling in fetal, aged, and Alzheimer hippocampus: a continuum of stress-related signaling. Neurochem Res 29: 1287-1297. doi: 10.1023/B:NERE.0000023615.89699.63
|
| [21] |
Smith MA, Nunomura A, Lee HG, et al. (2005) Chronological primacy of oxidative stress in Alzheimer disease. Neurobiol Aging 26: 579-580; discussion 587-595. doi: 10.1016/j.neurobiolaging.2004.09.021
|
| [22] |
Overmyer M, Helisalmi S, Soininen H, et al. (1999) Reactive microglia in aging and dementia: an immunohistochemical study of postmortem human brain tissue. Acta Neuropathol (Berl) 97: 383-392. doi: 10.1007/s004010051002
|
| [23] |
Currais A (2015) Ageing and inflammation - A central role for mitochondria in brain health and disease. Ageing Res Rev 21: 30-42. doi: 10.1016/j.arr.2015.02.001
|
| [24] | Mielke MM, Vemuri P, Rocca WA (2014) Clinical epidemiology of Alzheimer's disease: assessing sex and gender differences. Clin Epidemiol 6: 37-48. |
| [25] |
Chapman RM, Mapstone M, Gardner MN, et al. (2011) Women have farther to fall: gender differences between normal elderly and Alzheimer's disease in verbal memory engender better detection of Alzheimer's disease in women. J Int Neuropsychol Soc 17: 654-662. doi: 10.1017/S1355617711000452
|
| [26] |
Green PS, Simpkins JW (2000) Neuroprotective effects of estrogens: potential mechanisms of action. Int J Dev Neurosci 18: 347-358. doi: 10.1016/S0736-5748(00)00017-4
|
| [27] |
Jones KJ, Brown TJ, Damaser M (2001) Neuroprotective effects of gonadal steroids on regenerating peripheral motoneurons. Brain Res Brain Res Rev 37: 372-382. doi: 10.1016/S0165-0173(01)00107-2
|
| [28] | Wise PM, Dubal DB, Wilson ME, et al. (2001) Minireview: neuroprotective effects of estrogen-new insights into mechanisms of action. Endocrinology 142: 969-973. |
| [29] |
Strittmatter WJ, Saunders AM, Schmechel D, et al. (1993) Apolipoprotein E: high-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc Natl Acad Sci U S A 90: 1977-1981. doi: 10.1073/pnas.90.5.1977
|
| [30] |
Saunders AM, Strittmatter WJ, Schmechel D, et al. (1993) Association of apolipoprotein E allele epsilon 4 with late-onset familial and sporadic Alzheimer's disease. Neurology 43: 1467-1472. doi: 10.1212/WNL.43.8.1467
|
| [31] |
Corder EH, Saunders AM, Strittmatter WJ, et al. (1993) Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer's disease in late onset families. Science 261: 921-923. doi: 10.1126/science.8346443
|
| [32] |
McGeer PL, Walker DG, Pitas RE, et al. (1997) Apolipoprotein E4 (ApoE4) but not ApoE3 or ApoE2 potentiates beta- amyloid protein activation of complement in vitro. Brain Res 749: 135-138. doi: 10.1016/S0006-8993(96)01324-8
|
| [33] |
Prince JA, Zetterberg H, Andreasen N, et al. (2004) APOE epsilon4 allele is associated with reduced cerebrospinal fluid levels of Abeta42. Neurology 62: 2116-2118. doi: 10.1212/01.WNL.0000128088.08695.05
|
| [34] |
Tiraboschi P, Hansen LA, Masliah E, et al. (2004) Impact of APOE genotype on neuropathologic and neurochemical markers of Alzheimer disease. Neurology 62: 1977-1983. doi: 10.1212/01.WNL.0000128091.92139.0F
|
| [35] |
Lovestone S, Anderton BH, Hartley C, et al. (1996) The intracellular fate of apolipoprotein E is tau dependent and apoe allele-specific. Neuroreport 7: 1005-1008. doi: 10.1097/00001756-199604100-00010
|
| [36] |
Tesseur I, Van Dorpe J, Spittaels K, et al. (2000) Expression of human apolipoprotein E4 in neurons causes hyperphosphorylation of protein tau in the brains of transgenic mice. Am J Pathol 156: 951-964. doi: 10.1016/S0002-9440(10)64963-2
|
| [37] | Arendt T, Schindler C, Bruckner MK, et al. (1997) Plastic neuronal remodeling is impaired in patients with Alzheimer's disease carrying apolipoprotein epsilon 4 allele. J Neurosci 17: 516-529. |
| [38] | Burkhardt MS, Foster JK, Laws SM, et al. (2004) Oestrogen replacement therapy may improve memory functioning in the absence of APOE epsilon4. J Alzheimers Dis 6: 221-228. |
| [39] |
Patterson CE, Todd SA, Passmore AP (2011) Effect of apolipoprotein E and butyrylcholinesterase genotypes on cognitive response to cholinesterase inhibitor treatment at different stages of Alzheimer's disease. Pharmacogenomics J 11: 444-450. doi: 10.1038/tpj.2010.61
|
| [40] |
Petersen RC, Thomas RG, Grundman M, et al. (2005) Vitamin E and donepezil for the treatment of mild cognitive impairment. N Engl J Med 352: 2379-2388. doi: 10.1056/NEJMoa050151
|
| [41] |
Bettens K, Sleegers K, Van Broeckhoven C (2013) Genetic insights in Alzheimer's disease. Lancet Neurol 12: 92-104. doi: 10.1016/S1474-4422(12)70259-4
|
| [42] |
Norton S, Matthews FE, Barnes DE, et al. (2014) Potential for primary prevention of Alzheimer's disease: an analysis of population-based data. Lancet Neurol 13: 788-794. doi: 10.1016/S1474-4422(14)70136-X
|
| [43] |
Van Den Heuvel C, Thornton E, Vink R (2007) Traumatic brain injury and Alzheimer's disease: a review. Prog Brain Res 161: 303-316. doi: 10.1016/S0079-6123(06)61021-2
|
| [44] |
Solomon A, Kareholt I, Ngandu T, et al. (2007) Serum cholesterol changes after midlife and late-life cognition: twenty-one-year follow-up study. Neurology 68: 751-756. doi: 10.1212/01.wnl.0000256368.57375.b7
|
| [45] | Chin-Chan M, Navarro-Yepes J, Quintanilla-Vega B (2015) Environmental pollutants as risk factors for neurodegenerative disorders: Alzheimer and Parkinson diseases. Front Cell Neurosci 9: 124. |
| [46] | Organization WH (2015) Global Strategy on Diet, Physical Activity and Health. |
| [47] |
Warburton DE, Nicol CW, Bredin SS (2006) Health benefits of physical activity: the evidence. CMAJ 174: 801-809. doi: 10.1503/cmaj.051351
|
| [48] |
Beckett MW, Ardern CI, Rotondi MA (2015) A meta-analysis of prospective studies on the role of physical activity and the prevention of Alzheimer's disease in older adults. BMC Geriatr 15: 9. doi: 10.1186/s12877-015-0007-2
|
| [49] |
Reiner M, Niermann C, Jekauc D, et al. (2013) Long-term health benefits of physical activity--a systematic review of longitudinal studies. BMC Public Health 13: 813. doi: 10.1186/1471-2458-13-813
|
| [50] | Angevaren M, Aufdemkampe G, Verhaar HJ, et al. (2008) Physical activity and enhanced fitness to improve cognitive function in older people without known cognitive impairment. Cochrane Database Syst Rev: CD005381. |
| [51] | Carter ND, Khan KM, McKay HA, et al. (2002) Community-based exercise program reduces risk factors for falls in 65- to 75-year-old women with osteoporosis: randomized controlled trial. CMAJ 167: 997-1004. |
| [52] |
Lynch NA, Ryan AS, Evans J, et al. (2007) Older elite football players have reduced cardiac and osteoporosis risk factors. Med Sci Sports Exerc 39: 1124-1130. doi: 10.1249/01.mss.0b013e3180557466
|
| [53] |
Vuori IM (2001) Dose-response of physical activity and low back pain, osteoarthritis, and osteoporosis. Med Sci Sports Exerc 33: S551-586; discussion 609-510. doi: 10.1097/00005768-200106001-00026
|
| [54] |
Tan ZS, Seshadri S, Beiser A, et al. (2005) Bone mineral density and the risk of Alzheimer disease. Arch Neurol 62: 107-111. doi: 10.1001/archneur.62.1.107
|
| [55] |
Zhang Y, Seshadri S, Ellison RC, et al. (2001) Bone mineral density and verbal memory impairment: Third National Health and Nutrition Examination Survey. Am J Epidemiol 154: 795-802. doi: 10.1093/aje/154.9.795
|
| [56] |
Yaffe K, Browner W, Cauley J, et al. (1999) Association between bone mineral density and cognitive decline in older women. J Am Geriatr Soc 47: 1176-1182. doi: 10.1111/j.1532-5415.1999.tb05196.x
|
| [57] |
Gunstad J, Paul RH, Cohen RA, et al. (2007) Elevated body mass index is associated with executive dysfunction in otherwise healthy adults. Compr Psychiatry 48: 57-61. doi: 10.1016/j.comppsych.2006.05.001
|
| [58] |
Gustafson D, Lissner L, Bengtsson C, et al. (2004) A 24-year follow-up of body mass index and cerebral atrophy. Neurology 63: 1876-1881. doi: 10.1212/01.WNL.0000141850.47773.5F
|
| [59] | Jagust W, Harvey D, Mungas D, et al. (2005) Central obesity and the aging brain. Arch Neurol 62: 1545-1548. |
| [60] | Kivipelto M, Ngandu T, Fratiglioni L, et al. (2005) Obesity and vascular risk factors at midlife and the risk of dementia and Alzheimer disease. Arch Neurol 62: 1556-1560. |
| [61] |
Whitmer RA, Gunderson EP, Barrett-Connor E, et al. (2005) Obesity in middle age and future risk of dementia: a 27 year longitudinal population based study. Bmj 330: 1360. doi: 10.1136/bmj.38446.466238.E0
|
| [62] | Sohrabi HR, Bates KA, Weinborn M, et al. (2015) Bone mineral density, adiposity, and cognitive functions. Front Aging Neurosci 7: 16. |
| [63] |
Koudinov AR, Berezov TT, Kumar A, et al. (1998) Alzheimer's amyloid beta interaction with normal human plasma high density lipoprotein: association with apolipoprotein and lipids. Clin Chim Acta 270: 75-84. doi: 10.1016/S0009-8981(97)00207-6
|
| [64] |
Koudinov AR, Koudinova NV, Kumar A, et al. (1996) Biochemical characterization of Alzheimer's soluble amyloid beta protein in human cerebrospinal fluid: Associations with high density lipoproteins. Biochem Biophys Res Commun 223: 592-597. doi: 10.1006/bbrc.1996.0940
|
| [65] | Eapen DJ, Kalra GL, Rifai L, et al. (2010) Raising HDL cholesterol in women. Int J Womens Health 1: 181-191. |
| [66] | Bates KA, Sohrabi HR, Rodrigues M, et al. (2009) Association of Cardiovascular Factors and Alzheimer's Disease Plasma Amyloid-beta Protein in Subjective Memory Complainers. J Alzheimers Dis 17: 305-318. |
| [67] |
Atzmon G, Gabriely I, Greiner W, et al. (2002) Plasma HDL levels highly correlate with cognitive function in exceptional longevity. J Gerontol A Biol Sci Med Sci 57: M712-715. doi: 10.1093/gerona/57.11.M712
|
| [68] |
Crichton GE, Elias MF, Davey A, et al. (2014) Higher HDL cholesterol is associated with better cognitive function: the Maine-Syracuse study. J Int Neuropsychol Soc 20: 961-970. doi: 10.1017/S1355617714000885
|
| [69] | Singh-Manoux A, Gimeno D, Kivimaki M, et al. (2008) Low HDL cholesterol is a risk factor for deficit and decline in memory in midlife. The Whitehall II study. Arterioscler Thromb Vasc Biol 28: 1557-1563. |
| [70] | Ward MA, Bendlin BB, McLaren DG, et al. (2010) Low HDL Cholesterol is Associated with Lower Gray Matter Volume in Cognitively Healthy Adults. Front Aging Neurosci 2. |
| [71] | Biessels GJ, Kappelle LJ (2005) Increased risk of Alzheimer's disease in Type II diabetes: insulin resistance of the brain or insulin-induced amyloid pathology? Biochem Soc Trans 33: 1041-1044. |
| [72] |
Gatto NM, Henderson VW, St John JA, et al. (2008) Metabolic syndrome and cognitive function in healthy middle-aged and older adults without diabetes. Neuropsychol Dev Cogn B Aging Neuropsychol Cogn 15: 627-641. doi: 10.1080/13825580802036936
|
| [73] |
Tomlinson DR, Gardiner NJ (2008) Glucose neurotoxicity. Nat Rev Neurosci 9: 36-45. doi: 10.1038/nrn2294
|
| [74] |
Craft S, Asthana S, Newcomer JW, et al. (1999) Enhancement of memory in Alzheimer disease with insulin and somatostatin, but not glucose. Arch Gen Psychiatry 56: 1135-1140. doi: 10.1001/archpsyc.56.12.1135
|
| [75] |
Craft S, Newcomer J, Kanne S, et al. (1996) Memory improvement following induced hyperinsulinemia in Alzheimer's disease. Neurobiol Aging 17: 123-130. doi: 10.1016/0197-4580(95)02002-0
|
| [76] |
Kulstad JJ, Green PS, Cook DG, et al. (2006) Differential modulation of plasma beta-amyloid by insulin in patients with Alzheimer disease. Neurology 66: 1506-1510. doi: 10.1212/01.wnl.0000216274.58185.09
|
| [77] |
Ibanez J, Izquierdo M, Arguelles I, et al. (2005) Twice-weekly progressive resistance training decreases abdominal fat and improves insulin sensitivity in older men with type 2 diabetes. Diabetes Care 28: 662-667. doi: 10.2337/diacare.28.3.662
|
| [78] |
Knowler WC, Barrett-Connor E, Fowler SE, et al. (2002) Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N Engl J Med 346: 393-403. doi: 10.1056/NEJMoa012512
|
| [79] |
Cotman CW, Berchtold NC, Christie LA (2007) Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci 30: 464-472. doi: 10.1016/j.tins.2007.06.011
|
| [80] | Kuipers SD, Bramham CR (2006) Brain-derived neurotrophic factor mechanisms and function in adult synaptic plasticity: new insights and implications for therapy. Curr Opin Drug Discov Devel 9: 580-586. |
| [81] |
Neeper SA, Gomez-Pinilla F, Choi J, et al. (1996) Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res 726: 49-56. doi: 10.1016/0006-8993(96)00273-9
|
| [82] |
Oliff HS, Berchtold NC, Isackson P, et al. (1998) Exercise-induced regulation of brain-derived neurotrophic factor (BDNF) transcripts in the rat hippocampus. Brain Res Mol Brain Res 61: 147-153. doi: 10.1016/S0169-328X(98)00222-8
|
| [83] |
Widenfalk J, Olson L, Thoren P (1999) Deprived of habitual running, rats downregulate BDNF and TrkB messages in the brain. Neurosci Res 34: 125-132. doi: 10.1016/S0168-0102(99)00051-6
|
| [84] |
Vaynman S, Ying Z, Gomez-Pinilla F (2004) Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur J Neurosci 20: 2580-2590. doi: 10.1111/j.1460-9568.2004.03720.x
|
| [85] |
Vaynman SS, Ying Z, Yin D, et al. (2006) Exercise differentially regulates synaptic proteins associated to the function of BDNF. Brain Res 1070: 124-130. doi: 10.1016/j.brainres.2005.11.062
|
| [86] |
Kolb EM, Rezende EL, Holness L, et al. (2013) Mice selectively bred for high voluntary wheel running have larger midbrains: support for the mosaic model of brain evolution. J Exp Biol 216: 515-523. doi: 10.1242/jeb.076000
|
| [87] | Carro E, Nunez A, Busiguina S, et al. (2000) Circulating insulin-like growth factor I mediates effects of exercise on the brain. J Neurosci 20: 2926-2933. |
| [88] |
Ding Q, Vaynman S, Akhavan M, et al. (2006) Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function. Neuroscience 140: 823-833. doi: 10.1016/j.neuroscience.2006.02.084
|
| [89] |
McCusker RH, McCrea K, Zunich S, et al. (2006) Insulin-like growth factor-I enhances the biological activity of brain-derived neurotrophic factor on cerebrocortical neurons. J Neuroimmunol 179: 186-190. doi: 10.1016/j.jneuroim.2006.06.014
|
| [90] |
Carro E, Torres-Aleman I (2004) The role of insulin and insulin-like growth factor I in the molecular and cellular mechanisms underlying the pathology of Alzheimer's disease. Eur J Pharmacol 490: 127-133. doi: 10.1016/j.ejphar.2004.02.050
|
| [91] |
Erickson KI, Raji CA, Lopez OL, et al. (2010) Physical activity predicts gray matter volume in late adulthood: the Cardiovascular Health Study. Neurology 75: 1415-1422. doi: 10.1212/WNL.0b013e3181f88359
|
| [92] |
Bugg JM, Head D (2011) Exercise moderates age-related atrophy of the medial temporal lobe. Neurobiol Aging 32: 506-514. doi: 10.1016/j.neurobiolaging.2009.03.008
|
| [93] | Boyle CP, Raji CA, Erickson KI, et al. (2015) Physical activity, body mass index, and brain atrophy in Alzheimer's disease. Neurobiol Aging 36 Suppl 1: S194-202. |
| [94] |
Rovio S, Spulber G, Nieminen LJ, et al. (2010) The effect of midlife physical activity on structural brain changes in the elderly. Neurobiol Aging 31: 1927-1936. doi: 10.1016/j.neurobiolaging.2008.10.007
|
| [95] |
Erickson KI, Voss MW, Prakash RS, et al. (2011) Exercise training increases size of hippocampus and improves memory. Proc Natl Acad Sci U S A 108: 3017-3022. doi: 10.1073/pnas.1015950108
|
| [96] |
Burns JM, Cronk BB, Anderson HS, et al. (2008) Cardiorespiratory fitness and brain atrophy in early Alzheimer disease. Neurology 71: 210-216. doi: 10.1212/01.wnl.0000317094.86209.cb
|
| [97] |
Honea RA, Thomas GP, Harsha A, et al. (2009) Cardiorespiratory fitness and preserved medial temporal lobe volume in Alzheimer disease. Alzheimer Dis Assoc Disord 23: 188-197. doi: 10.1097/WAD.0b013e31819cb8a2
|
| [98] |
van Praag H, Shubert T, Zhao C, et al. (2005) Exercise enhances learning and hippocampal neurogenesis in aged mice. J Neurosci 25: 8680-8685. doi: 10.1523/JNEUROSCI.1731-05.2005
|
| [99] |
Albeck DS, Sano K, Prewitt GE, et al. (2006) Mild forced treadmill exercise enhances spatial learning in the aged rat. Behav Brain Res 168: 345-348. doi: 10.1016/j.bbr.2005.11.008
|
| [100] |
Kramer AF, Erickson KI (2007) Capitalizing on cortical plasticity: influence of physical activity on cognition and brain function. Trends Cogn Sci 11: 342-348. doi: 10.1016/j.tics.2007.06.009
|
| [101] |
Heyn P, Abreu BC, Ottenbacher KJ (2004) The effects of exercise training on elderly persons with cognitive impairment and dementia: a meta-analysis. Arch Phys Med Rehabil 85: 1694-1704. doi: 10.1016/j.apmr.2004.03.019
|
| [102] |
Colcombe S, Kramer AF (2003) Fitness effects on the cognitive function of older adults: a meta-analytic study. Psychol Sci 14: 125-130. doi: 10.1111/1467-9280.t01-1-01430
|
| [103] |
Stones MJ, Kozma A (1989) Age, exercise, and coding performance. Psychol Aging 4: 190-194. doi: 10.1037/0882-7974.4.2.190
|
| [104] |
Kamijo K, Takeda Y, Hillman CH (2011) The relation of physical activity to functional connectivity between brain regions. Clin Neurophysiol 122: 81-89. doi: 10.1016/j.clinph.2010.06.007
|
| [105] | Burdette JH, Laurienti PJ, Espeland MA, et al. (2010) Using network science to evaluate exercise-associated brain changes in older adults. Front Aging Neurosci 2: 23. |
| [106] | Voss MW, Prakash RS, Erickson KI, et al. (2010) Plasticity of brain networks in a randomized intervention trial of exercise training in older adults. Front Aging Neurosci 2. |
| [107] |
Strawbridge WJ, Deleger S, Roberts RE, et al. (2002) Physical activity reduces the risk of subsequent depression for older adults. Am J Epidemiol 156: 328-334. doi: 10.1093/aje/kwf047
|
| [108] |
Lampinen P, Heikkinen E (2003) Reduced mobility and physical activity as predictors of depressive symptoms among community-dwelling older adults: an eight-year follow-up study. Aging Clin Exp Res 15: 205-211. doi: 10.1007/BF03324501
|
| [109] |
Lee Y, Park K (2008) Does physical activity moderate the association between depressive symptoms and disability in older adults? Int J Geriatr Psychiatry 23: 249-256. doi: 10.1002/gps.1870
|
| [110] |
Yoshiuchi K, Nakahara R, Kumano H, et al. (2006) Yearlong physical activity and depressive symptoms in older Japanese adults: cross-sectional data from the Nakanojo study. Am J Geriatr Psychiatry 14: 621-624. doi: 10.1097/01.JGP.0000200602.70504.9c
|
| [111] |
Salguero A, Martinez-Garcia R, Molinero O, et al. (2011) Physical activity, quality of life and symptoms of depression in community-dwelling and institutionalized older adults. Arch Gerontol Geriatr 53: 152-157. doi: 10.1016/j.archger.2010.10.005
|
| [112] |
Lee H, Lee JA, Brar JS, et al. (2014) Physical activity and depressive symptoms in older adults. Geriatr Nurs 35: 37-41. doi: 10.1016/j.gerinurse.2013.09.005
|
| [113] |
Maki Y, Ura C, Yamaguchi T, et al. (2012) Effects of intervention using a community-based walking program for prevention of mental decline: a randomized controlled trial. J Am Geriatr Soc 60: 505-510. doi: 10.1111/j.1532-5415.2011.03838.x
|
| [114] |
Penninx BW, Rejeski WJ, Pandya J, et al. (2002) Exercise and depressive symptoms: a comparison of aerobic and resistance exercise effects on emotional and physical function in older persons with high and low depressive symptomatology. J Gerontol B Psychol Sci Soc Sci 57: P124-132. doi: 10.1093/geronb/57.2.P124
|
| [115] |
Teixeira CM, Vasconcelos-Raposo J, Fernandes HM, et al. (2013) Physical Activity, Depression and Anxiety Among the Elderly. Social Indicators Research 113: 307-318. doi: 10.1007/s11205-012-0094-9
|
| [116] |
Lazarov O, Robinson J, Tang YP, et al. (2005) Environmental enrichment reduces Abeta levels and amyloid deposition in transgenic mice. Cell 120: 701-713. doi: 10.1016/j.cell.2005.01.015
|
| [117] |
Adlard PA, Perreau VM, Pop V, et al. (2005) Voluntary exercise decreases amyloid load in a transgenic model of Alzheimer's disease. J Neurosci 25: 4217-4221. doi: 10.1523/JNEUROSCI.0496-05.2005
|
| [118] |
Kemppainen NM, Aalto S, Wilson IA, et al. (2006) Voxel-based analysis of PET amyloid ligand [11C]PIB uptake in Alzheimer disease. Neurology 67: 1575-1580. doi: 10.1212/01.wnl.0000240117.55680.0a
|
| [119] |
Klunk WE, Engler H, Nordberg A, et al. (2004) Imaging brain amyloid in Alzheimer's disease with Pittsburgh Compound-B. Ann Neurol 55: 306-319. doi: 10.1002/ana.20009
|
| [120] |
Klunk WE, Lopresti BJ, Ikonomovic MD, et al. (2005) Binding of the positron emission tomography tracer Pittsburgh compound-B reflects the amount of amyloid-beta in Alzheimer's disease brain but not in transgenic mouse brain. J Neurosci 25: 10598-10606. doi: 10.1523/JNEUROSCI.2990-05.2005
|
| [121] |
Liang KY, Mintun MA, Fagan AM, et al. (2010) Exercise and Alzheimer's disease biomarkers in cognitively normal older adults. Ann Neurol 68: 311-318. doi: 10.1002/ana.22096
|
| [122] |
Vemuri P, Lesnick TG, Przybelski SA, et al. (2012) Effect of lifestyle activities on Alzheimer disease biomarkers and cognition. Ann Neurol 72: 730-738. doi: 10.1002/ana.23665
|
| [123] |
Brown BM, Peiffer JJ, Taddei K, et al. (2013) Physical activity and amyloid-beta plasma and brain levels: results from the Australian Imaging, Biomarkers and Lifestyle Study of Ageing. Mol Psychiatry 18: 875-881. doi: 10.1038/mp.2012.107
|
| [124] |
Okonkwo OC, Schultz SA, Oh JM, et al. (2014) Physical activity attenuates age-related biomarker alterations in preclinical AD. Neurology 83: 1753-1760. doi: 10.1212/WNL.0000000000000964
|
| [125] |
Bennet AM, Di Angelantonio E, Ye Z, et al. (2007) Association of apolipoprotein E genotypes with lipid levels and coronary risk. Jama 298: 1300-1311. doi: 10.1001/jama.298.11.1300
|
| [126] | Mahley RW, Nathan BP, Pitas RE (1996) Apolipoprotein E. Structure, function, and possible roles in Alzheimer's disease. Ann N Y Acad Sci 777: 139-145. |
| [127] |
Weisgraber KH (1994) Apolipoprotein E: structure-function relationships. Adv Protein Chem 45: 249-302. doi: 10.1016/S0065-3233(08)60642-7
|
| [128] | Weisgraber KH, Mahley RW (1996) Human apolipoprotein E: the Alzheimer's disease connection. Faseb J 10: 1485-1494. |
| [129] |
Yaffe K, Haan M, Byers A, et al. (2000) Estrogen use, APOE, and cognitive decline: evidence of gene-environment interaction. Neurology 54: 1949-1954. doi: 10.1212/WNL.54.10.1949
|
| [130] |
Szekely CA, Breitner JC, Fitzpatrick AL, et al. (2008) NSAID use and dementia risk in the Cardiovascular Health Study: role of APOE and NSAID type. Neurology 70: 17-24. doi: 10.1212/01.wnl.0000284596.95156.48
|
| [131] |
Qiu C, Winblad B, Fastbom J, et al. (2003) Combined effects of APOE genotype, blood pressure, and antihypertensive drug use on incident AD. Neurology 61: 655-660. doi: 10.1212/WNL.61.5.655
|
| [132] |
Rovio S, Kareholt I, Helkala EL, et al. (2005) Leisure-time physical activity at midlife and the risk of dementia and Alzheimer's disease. Lancet Neurol 4: 705-711. doi: 10.1016/S1474-4422(05)70198-8
|
| [133] |
Deeny SP, Poeppel D, Zimmerman JB, et al. (2008) Exercise, APOE, and working memory: MEG and behavioral evidence for benefit of exercise in epsilon4 carriers. Biol Psychol 78: 179-187. doi: 10.1016/j.biopsycho.2008.02.007
|
| [134] |
Etnier JL, Caselli RJ, Reiman EM, et al. (2007) Cognitive performance in older women relative to ApoE-epsilon4 genotype and aerobic fitness. Med Sci Sports Exerc 39: 199-207. doi: 10.1249/01.mss.0000239399.85955.5e
|
| [135] |
Kivipelto M, Rovio S, Ngandu T, et al. (2008) Apolipoprotein E epsilon4 magnifies lifestyle risks for dementia: a population-based study. J Cell Mol Med 12: 2762-2771. doi: 10.1111/j.1582-4934.2008.00296.x
|
| [136] | Niti M, Yap KB, Kua EH, et al. (2008) Physical, social and productive leisure activities, cognitive decline and interaction with APOE-epsilon 4 genotype in Chinese older adults. Int Psychogeriatr 20: 237-251. |
| [137] | Schuit AJ, Feskens EJ, Launer LJ, et al. (2001) Physical activity and cognitive decline, the role of the apolipoprotein e4 allele. Med Sci Sports Exerc 33: 772-777. |
| [138] | Yang SY, Weng PH, Chen JH, et al. (2014) Leisure activities, apolipoprotein E e4 status, and the risk of dementia. J Formos Med Assoc. |
| [139] |
Luck T, Riedel-Heller SG, Luppa M, et al. (2014) Apolipoprotein E epsilon 4 genotype and a physically active lifestyle in late life: analysis of gene-environment interaction for the risk of dementia and Alzheimer's disease dementia. Psychol Med 44: 1319-1329. doi: 10.1017/S0033291713001918
|
| [140] |
Podewils LJ, Guallar E, Kuller LH, et al. (2005) Physical Activity, APOE Genotype, and Dementia Risk: Findings from the Cardiovascular Health Cognition Study. Am J Epidemiol 161: 639-651. doi: 10.1093/aje/kwi092
|
| [141] |
Lindsay J, Laurin D, Verreault R, et al. (2002) Risk factors for Alzheimer's disease: a prospective analysis from the Canadian Study of Health and Aging. Am J Epidemiol 156: 445-453. doi: 10.1093/aje/kwf074
|
| [142] |
Sabia S, Kivimaki M, Kumari M, et al. (2010) Effect of Apolipoprotein E epsilon4 on the association between health behaviors and cognitive function in late midlife. Mol Neurodegener 5: 23. doi: 10.1186/1750-1326-5-23
|
| [143] |
Taaffe DR, Irie F, Masaki KH, et al. (2008) Physical activity, physical function, and incident dementia in elderly men: the Honolulu-Asia Aging Study. J Gerontol A Biol Sci Med Sci 63: 529-535. doi: 10.1093/gerona/63.5.529
|
| [144] | Smith JC, Nielson KA, Woodard JL, et al. (2014) Physical activity reduces hippocampal atrophy in elders at genetic risk for Alzheimer's disease. Front Aging Neurosci 6: 61. |
| [145] |
Head D, Bugg JM, Goate AM, et al. (2012) Exercise Engagement as a Moderator of the Effects of APOE Genotype on Amyloid Deposition. Arch Neurol 69: 636-643. doi: 10.1001/archneurol.2011.845
|
| [146] |
Nichol K, Deeny SP, Seif J, et al. (2009) Exercise improves cognition and hippocampal plasticity in APOE epsilon4 mice. Alzheimers Dement 5: 287-294. doi: 10.1016/j.jalz.2009.02.006
|
| [147] | Buttini M, Yu GQ, Shockley K, et al. (2002) Modulation of Alzheimer-like synaptic and cholinergic deficits in transgenic mice by human apolipoprotein E depends on isoform, aging, and overexpression of amyloid beta peptides but not on plaque formation. J Neurosci 22: 10539-10548. |
| [148] |
Allen SJ, MacGowan SH, Tyler S, et al. (1997) Reduced cholinergic function in normal and Alzheimer's disease brain is associated with apolipoprotein E4 genotype. Neurosci Lett 239: 33-36. doi: 10.1016/S0304-3940(97)00872-0
|
| [149] | Scarmeas N, Habeck C, Anderson KE, et al. (2004) Altered PET functional brain responses in cognitively intact elderly persons at risk for Alzheimer disease (carriers of the epsilon4 allele). Am J Geriatr Psychiatry 12: 596-605. |
| [150] | Rimajova M, Lenzo NP, Wu J-S, et al. (2007) Fluoro-2-deoxy-D-glucose (FDG)-PET in APOEε4 carriers in the Australian population. Journal of Alzheimer's disease 13: 137-146. |
| [151] |
Mosconi L, De Santi S, Brys M, et al. (2008) Hypometabolism and altered cerebrospinal fluid markers in normal apolipoprotein E E4 carriers with subjective memory complaints. Biol Psychiatry 63: 609-618. doi: 10.1016/j.biopsych.2007.05.030
|
| [152] | Hone E, Martins IJ, Fonte J, et al. (2003) Apolipoprotein E influences amyloid-beta clearance from the murine periphery. J Alzheimers Dis 5: 1-8. |
| [153] | Hone E, Martins IJ, Jeoung M, et al. (2005) Alzheimer's disease amyloid-beta peptide modulates apolipoprotein E isoform specific receptor binding. J Alzheimers Dis 7: 303-314. |
| [154] |
Mazur-Kolecka B, Frackowiak J, Kowal D, et al. (2002) Oxidative protein damage in cells engaged in beta-amyloidosis is related to apoE genotype. Neuroreport 13: 465-468. doi: 10.1097/00001756-200203250-00021
|
| [155] |
Overmyer M, Helisalmi S, Soininen H, et al. (1999) Astrogliosis and the ApoE genotype. an immunohistochemical study of postmortem human brain tissue. Dement Geriatr Cogn Disord 10: 252-257. doi: 10.1159/000017128
|
| [156] |
Taddei K, Clarnette R, Gandy SE, et al. (1997) Increased plasma apolipoprotein E (apoE) levels in Alzheimer's disease. Neurosci Lett 223: 29-32. doi: 10.1016/S0304-3940(97)13394-8
|
| [157] |
Laws SM, Hone E, Gandy S, et al. (2003) Expanding the association between the APOE gene and the risk of Alzheimer's disease: possible roles for APOE promoter polymorphisms and alterations in APOE transcription. J Neurochem 84: 1215-1236. doi: 10.1046/j.1471-4159.2003.01615.x
|
| [158] |
Laws SM, Hone E, Taddei K, et al. (2002) Variation at the APOE -491 promoter locus is associated with altered brain levels of apolipoprotein E. Mol Psychiatry 7: 886-890. doi: 10.1038/sj.mp.4001097
|
| [159] | Casadei VM, Ferri C, Veglia F, et al. (1999) APOE-491 promoter polymorphism is a risk factor for late-onset Alzheimer's disease. Neurology 53: 1888-1889. |
| [160] |
Bueller JA, Aftab M, Sen S, et al. (2006) BDNF Val66Met allele is associated with reduced hippocampal volume in healthy subjects. Biol Psychiatry 59: 812-815. doi: 10.1016/j.biopsych.2005.09.022
|
| [161] |
Lim YY, Villemagne VL, Laws SM, et al. (2014) Effect of BDNF Val66Met on memory decline and hippocampal atrophy in prodromal Alzheimer's disease: a preliminary study. PLoS One 9: e86498. doi: 10.1371/journal.pone.0086498
|
| [162] |
Feher A, Juhasz A, Rimanoczy A, et al. (2009) Association between BDNF Val66Met polymorphism and Alzheimer disease, dementia with Lewy bodies, and Pick disease. Alzheimer Dis Assoc Disord 23: 224-228. doi: 10.1097/WAD.0b013e318199dd7d
|
| [163] |
Brown BM, Bourgeat P, Peiffer JJ, et al. (2014) Influence of BDNF Val66Met on the relationship between physical activity and brain volume. Neurology 83: 1345-1352. doi: 10.1212/WNL.0000000000000867
|
| [164] |
Teng HK, Teng KK, Lee R, et al. (2005) ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin. J Neurosci 25: 5455-5463. doi: 10.1523/JNEUROSCI.5123-04.2005
|
| [165] |
Zhang Y, Wang B, Wan H, et al. (2013) Meta-analysis of the insulin degrading enzyme polymorphisms and susceptibility to Alzheimer's disease. Neurosci Lett 541: 132-137. doi: 10.1016/j.neulet.2013.01.051
|
| [166] | Vargas T, Martinez-Garcia A, Antequera D, et al. (2011) IGF-I gene variability is associated with an increased risk for AD. Neurobiol Aging 32: 556 e553-511. |
| [167] |
Braskie MN, Boyle CP, Rajagopalan P, et al. (2014) Physical activity, inflammation, and volume of the aging brain. Neuroscience 273: 199-209. doi: 10.1016/j.neuroscience.2014.05.005
|
| [168] | Licastro F, Porcellini E, Caruso C, et al. (2006) Genetic risk profiles for Alzheimer's disease: Integration of APOE genotype and variants that up-regulate inflammation. Neurobiol Aging. |
| [169] |
Culpan D, MacGowan SH, Ford JM, et al. (2003) Tumour necrosis factor-alpha gene polymorphisms and Alzheimer's disease. Neurosci Lett 350: 61-65. doi: 10.1016/S0304-3940(03)00854-1
|
| [170] |
Du Y, Dodel RC, Eastwood BJ, et al. (2000) Association of an interleukin 1 alpha polymorphism with Alzheimer's disease. Neurology 55: 480-483. doi: 10.1212/WNL.55.4.480
|
| [171] |
Grimaldi LM, Casadei VM, Ferri C, et al. (2000) Association of early-onset Alzheimer's disease with an interleukin-1alpha gene polymorphism. Ann Neurol 47: 361-365. doi: 10.1002/1531-8249(200003)47:3<361::AID-ANA12>3.0.CO;2-N
|
| [172] |
Hedley R, Hallmayer J, Groth DM, et al. (2002) Association of interleukin-1 polymorphisms with Alzheimer's disease in Australia. Ann Neurol 51: 795-797. doi: 10.1002/ana.10196
|
| [173] |
Koivisto AM, Helisalmi S, Pihlajamaki J, et al. (2005) Interleukin-6 promoter polymorphism and late-onset Alzheimer's disease in the Finnish population. J Neurogenet 19: 155-161. doi: 10.1080/01677060600569721
|
| [174] |
Laws SM, Perneczky R, Wagenpfeil S, et al. (2005) TNF polymorphisms in Alzheimer disease and functional implications on CSF beta-amyloid levels. Hum Mutat 26: 29-35. doi: 10.1002/humu.20180
|
| [175] |
Lio D, Annoni G, Licastro F, et al. (2006) Tumor necrosis factor-alpha -308A/G polymorphism is associated with age at onset of Alzheimer's disease. Mech Ageing Dev 127: 567-571. doi: 10.1016/j.mad.2006.01.015
|
| [176] |
Nicoll JA, Mrak RE, Graham DI, et al. (2000) Association of interleukin-1 gene polymorphisms with Alzheimer's disease. Ann Neurol 47: 365-368. doi: 10.1002/1531-8249(200003)47:3<365::AID-ANA13>3.0.CO;2-G
|
| [177] |
Sciacca FL, Ferri C, Licastro F, et al. (2003) Interleukin-1B polymorphism is associated with age at onset of Alzheimer's disease. Neurobiol Aging 24: 927-931. doi: 10.1016/S0197-4580(03)00011-3
|
| [178] |
Shibata N, Ohnuma T, Takahashi T, et al. (2002) Effect of IL-6 polymorphism on risk of Alzheimer disease: genotype-phenotype association study in Japanese cases. Am J Med Genet 114: 436-439. doi: 10.1002/ajmg.10417
|
| [179] |
Ferencz B, Laukka EJ, Welmer AK, et al. (2014) The benefits of staying active in old age: physical activity counteracts the negative influence of PICALM, BIN1, and CLU risk alleles on episodic memory functioning. Psychol Aging 29: 440-449. doi: 10.1037/a0035465
|
| [180] | Holler CJ, Davis PR, Beckett TL, et al. (2014) Bridging integrator 1 (BIN1) protein expression increases in the Alzheimer's disease brain and correlates with neurofibrillary tangle pathology. J Alzheimers Dis 42: 1221-1227. |
| [181] | Xu W, Tan L, Yu JT (2014) The Role of PICALM in Alzheimer's Disease. Mol Neurobiol. |
| [182] |
Herring A, Donath A, Yarmolenko M, et al. (2012) Exercise during pregnancy mitigates Alzheimer-like pathology in mouse offspring. Faseb J 26: 117-128. doi: 10.1096/fj.11-193193
|
| [183] |
Parnpiansil P, Jutapakdeegul N, Chentanez T, et al. (2003) Exercise during pregnancy increases hippocampal brain-derived neurotrophic factor mRNA expression and spatial learning in neonatal rat pup. Neurosci Lett 352: 45-48. doi: 10.1016/j.neulet.2003.08.023
|
| [184] | Horsburgh S, Robson-Ansley P, Adams R, et al. (2015) Exercise and inflammation-related epigenetic modifications: focus on DNA methylation. Exerc Immunol Rev 21: 26-41. |
| [185] |
Kaliman P, Parrizas M, Lalanza JF, et al. (2011) Neurophysiological and epigenetic effects of physical exercise on the aging process. Ageing Res Rev 10: 475-486. doi: 10.1016/j.arr.2011.05.002
|
| [186] | Ntanasis-Stathopoulos J, Tzanninis JG, Philippou A, et al. (2013) Epigenetic regulation on gene expression induced by physical exercise. J Musculoskelet Neuronal Interact 13: 133-146. |
| [187] | Hess NCL, Dieberg G, McFarlane JR, et al. (2014) The effect of exercise intervention on cognitive performance in persons at risk of, or with, dementia: A systematic review and meta-analysis. Healthy Aging Research 3: 1-10. |
| [188] |
Holthoff VA, Marschner K, Scharf M, et al. (2015) Effects of physical activity training in patients with Alzheimer's dementia: results of a pilot RCT study. PLoS One 10: e0121478. doi: 10.1371/journal.pone.0121478
|
| [189] | Etnier JL, Labban JD, Karper WB, et al. (2015) Innovative Research Design Exploring the Effects of Physical Activity and Genetics on Cognitive Performance in Community-Based Older Adults. J Aging Phys Act. |
| [190] |
Lautenschlager NT, Cox KL, Flicker L, et al. (2008) Effect of physical activity on cognitive function in older adults at risk for Alzheimer disease: a randomized trial. JAMA 300: 1027-1037. doi: 10.1001/jama.300.9.1027
|
| [191] |
Buchman AS, Boyle PA, Yu L, et al. (2012) Total daily physical activity and the risk of AD and cognitive decline in older adults. Neurology 78: 1323-1329. doi: 10.1212/WNL.0b013e3182535d35
|
| [192] | Zlatar ZZ, McGregor KM, Towler S, et al. (2015) Self-reported physical activity and objective aerobic fitness: differential associations with gray matter density in healthy aging. Front Aging Neurosci 7: 5. |
| [193] |
Akbaraly TN, Portet F, Fustinoni S, et al. (2009) Leisure activities and the risk of dementia in the elderly: results from the Three-City Study. Neurology 73: 854-861. doi: 10.1212/WNL.0b013e3181b7849b
|
| [194] |
Scarmeas N, Levy G, Tang MX, et al. (2001) Influence of leisure activity on the incidence of Alzheimer's disease. Neurology 57: 2236-2242. doi: 10.1212/WNL.57.12.2236
|
| [195] |
Schooler C, Mulatu MS (2001) The reciprocal effects of leisure time activities and intellectual functioning in older people: a longitudinal analysis. Psychol Aging 16: 466-482. doi: 10.1037/0882-7974.16.3.466
|
| [196] |
Verghese J, Lipton RB, Katz MJ, et al. (2003) Leisure activities and the risk of dementia in the elderly. N Engl J Med 348: 2508-2516. doi: 10.1056/NEJMoa022252
|
| [197] |
Barnes LL, Mendes de Leon CF, Wilson RS, et al. (2004) Social resources and cognitive decline in a population of older African Americans and whites. Neurology 63: 2322-2326. doi: 10.1212/01.WNL.0000147473.04043.B3
|
| [198] |
Bennett DA, Schneider JA, Tang Y, et al. (2006) The effect of social networks on the relation between Alzheimer's disease pathology and level of cognitive function in old people: a longitudinal cohort study. Lancet Neurol 5: 406-412. doi: 10.1016/S1474-4422(06)70417-3
|
| [199] |
Trost SG, Owen N, Bauman AE, et al. (2002) Correlates of adults' participation in physical activity: review and update. Med Sci Sports Exerc 34: 1996-2001. doi: 10.1097/00005768-200212000-00020
|
| [200] |
Dergance JM, Calmbach WL, Dhanda R, et al. (2003) Barriers to and benefits of leisure time physical activity in the elderly: differences across cultures. J Am Geriatr Soc 51: 863-868. doi: 10.1046/j.1365-2389.2003.51271.x
|
| [201] |
Kempermann G, Gast D, Gage FH (2002) Neuroplasticity in old age: sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann Neurol 52: 135-143. doi: 10.1002/ana.10262
|
| [202] |
Hannan AJ (2014) Environmental enrichment and brain repair: harnessing the therapeutic effects of cognitive stimulation and physical activity to enhance experience-dependent plasticity. Neuropathol Appl Neurobiol 40: 13-25. doi: 10.1111/nan.12102
|
| [203] |
Ball K, Berch DB, Helmers KF, et al. (2002) Effects of cognitive training interventions with older adults: a randomized controlled trial. Jama 288: 2271-2281. doi: 10.1001/jama.288.18.2271
|
| [204] | Calero MD, Navarro E (2007) Cognitive plasticity as a modulating variable on the effects of memory training in elderly persons. Arch Clin Neuropsychol 22: 63-72. |
| [205] |
Engvig A, Fjell AM, Westlye LT, et al. (2010) Effects of memory training on cortical thickness in the elderly. Neuroimage 52: 1667-1676. doi: 10.1016/j.neuroimage.2010.05.041
|
| [206] |
Rozzini L, Costardi D, Chilovi BV, et al. (2007) Efficacy of cognitive rehabilitation in patients with mild cognitive impairment treated with cholinesterase inhibitors. Int J Geriatr Psychiatry 22: 356-360. doi: 10.1002/gps.1681
|
| [207] |
Zanetti O, Binetti G, Magni E, et al. (1997) Procedural memory stimulation in Alzheimer's disease: impact of a training programme. Acta Neurol Scand 95: 152-157. doi: 10.1111/j.1600-0404.1997.tb00087.x
|
| [208] |
Zanetti O, Zanieri G, Di Giovanni G, et al. (2001) Effectiveness of procedural memory stimulation in mild Alzheimer's disease patients: A controlled study. Neuropsychol Rehabil 11: 263-272. doi: 10.1080/09602010042000088
|
| [209] |
Wilson RS, Mendes De Leon CF, Barnes LL, et al. (2002) Participation in cognitively stimulating activities and risk of incident Alzheimer disease. Jama 287: 742-748. doi: 10.1001/jama.287.6.742
|
| [210] |
Wilson RS, Scherr PA, Schneider JA, et al. (2007) Relation of cognitive activity to risk of developing Alzheimer disease. Neurology 69: 1911-1920. doi: 10.1212/01.wnl.0000271087.67782.cb
|
| [211] |
Shah T, Verdile G, Sohrabi H, et al. (2014) A combination of physical activity and computerized brain training improves verbal memory and increases cerebral glucose metabolism in the elderly. Transl Psychiatry 4: e487. doi: 10.1038/tp.2014.122
|
| [212] | Gates NJ, Sachdev P (2014) Is cognitive training an effective treatment for preclinical and early Alzheimer's disease? J Alzheimers Dis 42 Suppl 4: S551-559. |
| [213] |
Ngandu T, Lehtisalo J, Solomon A, et al. (2015) A 2 year multidomain intervention of diet, exercise, cognitive training, and vascular risk monitoring versus control to prevent cognitive decline in at-risk elderly people (FINGER): a randomised controlled trial. Lancet 385: 2255-2263. doi: 10.1016/S0140-6736(15)60461-5
|
| [214] |
Ellis KA, Bush AI, Darby D, et al. (2009) The Australian Imaging, Biomarkers and Lifestyle (AIBL) study of aging: methodology and baseline characteristics of 1112 individuals recruited for a longitudinal study of Alzheimer's disease. Int Psychogeriatr 21: 672-687. doi: 10.1017/S1041610209009405
|
| [215] |
Mueller SG, Weiner MW, Thal LJ, et al. (2005) Ways toward an early diagnosis in Alzheimer's disease: the Alzheimer's Disease Neuroimaging Initiative (ADNI). Alzheimers Dement 1: 55-66. doi: 10.1016/j.jalz.2005.06.003
|
| [216] |
Bateman RJ, Xiong C, Benzinger TL, et al. (2012) Clinical and biomarker changes in dominantly inherited Alzheimer's disease. N Engl J Med 367: 795-804. doi: 10.1056/NEJMoa1202753
|
| [217] |
Raber J, Wong D, Buttini M, et al. (1998) Isoform-specific effects of human apolipoprotein E on brain function revealed in ApoE knockout mice: increased susceptibility of females. Proc Natl Acad Sci U S A 95: 10914-10919. doi: 10.1073/pnas.95.18.10914
|
| [218] |
Ungar L, Altmann A, Greicius MD (2014) Apolipoprotein E, gender, and Alzheimer's disease: an overlooked, but potent and promising interaction. Brain Imaging Behav 8: 262-273. doi: 10.1007/s11682-013-9272-x
|
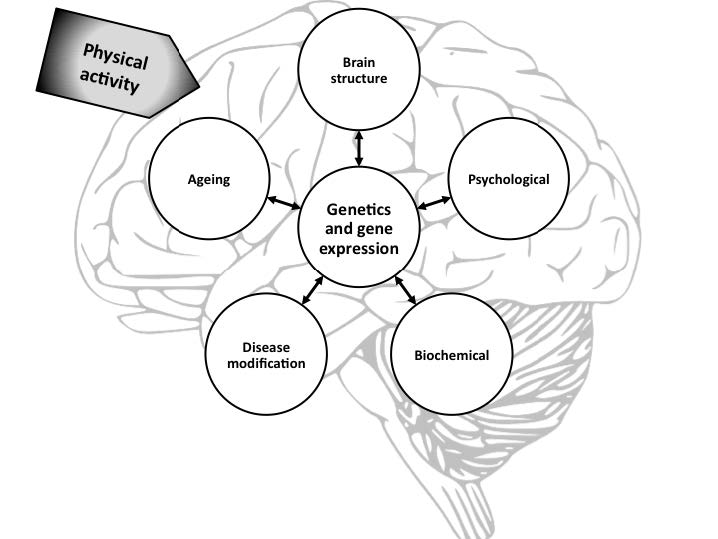
Figures(1) / Tables(1)

Kristyn Alissa Bates. Gene-environment interactions in considering physical activity for the prevention of dementia[J]. AIMS Molecular Science, 2015, 2(3): 359-381. doi: 10.3934/molsci.2015.3.359







 DownLoad:
DownLoad: