Citation: Isabel Saez, David Vilchez. Protein clearance mechanisms and their demise in age-related neurodegenerative diseases[J]. AIMS Molecular Science, 2015, 1(1): 1-21. doi: 10.3934/molsci.2015.1.1

| [1] |
Powers ET, Morimoto RI, Dillin A, et al. (2009) Biological and chemical approaches to diseases of proteostasis deficiency. Annu Rev Biochem 78: 959-991. doi: 10.1146/annurev.biochem.052308.114844

|
| [2] |
Hartl FU, Bracher A, Hayer-Hartl M (2011) Molecular chaperones in protein folding and proteostasis. Nature 475: 324-332. doi: 10.1038/nature10317
|
| [3] |
Balch WE, Morimoto RI, Dillin A, et al. (2008) Adapting proteostasis for disease intervention. Science 319: 916-919. doi: 10.1126/science.1141448
|
| [4] |
Tanaka K (2013) The proteasome: from basic mechanisms to emerging roles. Keio J Med 62: 1-12. doi: 10.2302/kjm.2012-0006-RE
|
| [5] | Wong E, Cuervo AM (2010) Integration of clearance mechanisms: the proteasome and autophagy. Cold Spring Harb Perspect Biol 2: a006734. |
| [6] |
Finley D (2009) Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu Rev Biochem 78: 477-513. doi: 10.1146/annurev.biochem.78.081507.101607
|
| [7] |
Lopez-Otin C, Blasco MA, Partridge L, et al. (2013) The hallmarks of aging. Cell 153: 1194-1217. doi: 10.1016/j.cell.2013.05.039
|
| [8] | Selkoe DJ (2011) Alzheimer's disease. Cold Spring Harb Perspect Biol 3. |
| [9] | Bosco DA, LaVoie MJ, Petsko GA, et al. (2011) Proteostasis and movement disorders: Parkinson's disease and amyotrophic lateral sclerosis. Cold Spring Harb Perspect Biol 3: a007500. |
| [10] | Finkbeiner S (2011) Huntington's Disease. Cold Spring Harb Perspect Biol 3. |
| [11] |
Schmidt M, Finley D (2014) Regulation of proteasome activity in health and disease. Biochim Biophys Acta 1843: 13-25. doi: 10.1016/j.bbamcr.2013.08.012
|
| [12] |
Tanaka K, Matsuda N (2014) Proteostasis and neurodegeneration: the roles of proteasomal degradation and autophagy. Biochim Biophys Acta 1843: 197-204. doi: 10.1016/j.bbamcr.2013.03.012
|
| [13] |
Vernace VA, Schmidt-Glenewinkel T, Figueiredo-Pereira ME (2007) Aging and regulated protein degradation: who has the UPPer hand? Aging Cell 6: 599-606. doi: 10.1111/j.1474-9726.2007.00329.x
|
| [14] | Taylor RC, Dillin A (2011) Aging as an event of proteostasis collapse. Cold Spring Harb Perspect Biol 3. |
| [15] |
Buckley SM, Aranda-Orgilles B, Strikoudis A, et al. (2012) Regulation of pluripotency and cellular reprogramming by the ubiquitin-proteasome system. Cell Stem Cell 11: 783-798. doi: 10.1016/j.stem.2012.09.011
|
| [16] |
Okita Y, Nakayama KI (2012) UPS delivers pluripotency. Cell Stem Cell 11: 728-730. doi: 10.1016/j.stem.2012.11.009
|
| [17] |
Vilchez D, Boyer L, Morantte I, et al. (2012) Increased proteasome activity in human embryonic stem cells is regulated by PSMD11. Nature 489: 304-308. doi: 10.1038/nature11468
|
| [18] |
Iwai K (2012) Diverse ubiquitin signaling in NF-kappaB activation. Trends Cell Biol 22: 355-364. doi: 10.1016/j.tcb.2012.04.001
|
| [19] |
Thrower JS, Hoffman L, Rechsteiner M, et al. (2000) Recognition of the polyubiquitin proteolytic signal. EMBO J 19: 94-102. doi: 10.1093/emboj/19.1.94
|
| [20] |
Kisselev AF, Goldberg AL (2005) Monitoring activity and inhibition of 26S proteasomes with fluorogenic peptide substrates. Methods Enzymol 398: 364-378. doi: 10.1016/S0076-6879(05)98030-0
|
| [21] | Jung T, Höhn A, Grune T (2013) The proteasome and the degradation of oxidized proteins: PartII - proteinoxidationandproteasomaldegradation. Redox Biology 2: 99-104. |
| [22] |
Demartino GN, Gillette TG (2007) Proteasomes: machines for all reasons. Cell 129: 659-662. doi: 10.1016/j.cell.2007.05.007
|
| [23] | Forster A, Masters EI, Whitby FG, et al. (2005) The 1.9 A structure of a proteasome-11S activator complex and implications for proteasome-PAN/PA700 interactions. Mol Cell 18: 589-599. |
| [24] | Dubiel W, Pratt G, Ferrell K, et al. (1992) Purification of an 11 S regulator of the multicatalytic protease. J Biol Chem 267: 22369-22377. |
| [25] | Ma CP, Slaughter CA, DeMartino GN (1992) Identification, purification, and characterization of a protein activator (PA28) of the 20 S proteasome (macropain). J Biol Chem 267: 10515-10523. |
| [26] |
Realini C, Jensen CC, Zhang Z, et al. (1997) Characterization of recombinant REGalpha, REGbeta, and REGgamma proteasome activators. J Biol Chem 272: 25483-25492. doi: 10.1074/jbc.272.41.25483
|
| [27] |
Sijts A, Sun Y, Janek K, et al. (2002) The role of the proteasome activator PA28 in MHC class I antigen processing. Mol Immunol 39: 165-169. doi: 10.1016/S0161-5890(02)00099-8
|
| [28] |
Li X, Amazit L, Long W, et al. (2007) Ubiquitin- and ATP-independent proteolytic turnover of p21 by the REGgamma-proteasome pathway. Mol Cell 26: 831-842. doi: 10.1016/j.molcel.2007.05.028
|
| [29] |
Blickwedehl J, Agarwal M, Seong C, et al. (2008) Role for proteasome activator PA200 and postglutamyl proteasome activity in genomic stability. Proc Natl Acad Sci U S A 105: 16165-16170. doi: 10.1073/pnas.0803145105
|
| [30] |
Schmidt M, Haas W, Crosas B, et al. (2005) The HEAT repeat protein Blm10 regulates the yeast proteasome by capping the core particle. Nat Struct Mol Biol 12: 294-303. doi: 10.1038/nsmb914
|
| [31] |
Egan D, Kim J, Shaw RJ, et al. (2011) The autophagy initiating kinase ULK1 is regulated via opposing phosphorylation by AMPK and mTOR. Autophagy 7: 643-644. doi: 10.4161/auto.7.6.15123
|
| [32] |
Rubinsztein DC, Marino G, Kroemer G (2011) Autophagy and aging. Cell 146: 682-695. doi: 10.1016/j.cell.2011.07.030
|
| [33] |
Ravikumar B, Sarkar S, Davies JE, et al. (2010) Regulation of mammalian autophagy in physiology and pathophysiology. Physiol Rev 90: 1383-1435. doi: 10.1152/physrev.00030.2009
|
| [34] |
Nixon RA (2013) The role of autophagy in neurodegenerative disease. Nat Med 19: 983-997. doi: 10.1038/nm.3232
|
| [35] |
Martinez-Vicente M, Cuervo AM (2007) Autophagy and neurodegeneration: when the cleaning crew goes on strike. Lancet Neurol 6: 352-361. doi: 10.1016/S1474-4422(07)70076-5
|
| [36] |
Cuervo AM (2004) Autophagy: in sickness and in health. Trends Cell Biol 14: 70-77. doi: 10.1016/j.tcb.2003.12.002
|
| [37] |
Mizushima N, Levine B, Cuervo AM, et al. (2008) Autophagy fights disease through cellular self-digestion. Nature 451: 1069-1075. doi: 10.1038/nature06639
|
| [38] |
Mizushima N, Komatsu M (2011) Autophagy: renovation of cells and tissues. Cell 147: 728-741. doi: 10.1016/j.cell.2011.10.026
|
| [39] |
Cuervo AM (2010) Chaperone-mediated autophagy: selectivity pays off. Trends Endocrinol Metab 21: 142-150. doi: 10.1016/j.tem.2009.10.003
|
| [40] |
He C, Klionsky DJ (2009) Regulation mechanisms and signaling pathways of autophagy. Annu Rev Genet 43: 67-93. doi: 10.1146/annurev-genet-102808-114910
|
| [41] |
Klionsky DJ, Cregg JM, Dunn WA, Jr., et al. (2003) A unified nomenclature for yeast autophagy-related genes. Dev Cell 5: 539-545. doi: 10.1016/S1534-5807(03)00296-X
|
| [42] |
Dubouloz F, Deloche O, Wanke V, et al. (2005) The TOR and EGO protein complexes orchestrate microautophagy in yeast. Mol Cell 19: 15-26. doi: 10.1016/j.molcel.2005.05.020
|
| [43] |
Marzella L, Ahlberg J, Glaumann H (1981) Autophagy, heterophagy, microautophagy and crinophagy as the means for intracellular degradation. Virchows Arch B Cell Pathol Incl Mol Pathol 36: 219-234. doi: 10.1007/BF02912068
|
| [44] | Mortimore GE, Lardeux BR, Adams CE (1988) Regulation of microautophagy and basal protein turnover in rat liver. Effects of short-term starvation. J Biol Chem 263: 2506-2512. |
| [45] |
Cuervo AM, Dice JF (1996) A receptor for the selective uptake and degradation of proteins by lysosomes. Science 273: 501-503. doi: 10.1126/science.273.5274.501
|
| [46] |
Rubinsztein DC, DiFiglia M, Heintz N, et al. (2005) Autophagy and its possible roles in nervous system diseases, damage and repair. Autophagy 1: 11-22. doi: 10.4161/auto.1.1.1513
|
| [47] |
Lee CK, Klopp RG, Weindruch R, et al. (1999) Gene expression profile of aging and its retardation by caloric restriction. Science 285: 1390-1393. doi: 10.1126/science.285.5432.1390
|
| [48] |
Ly DH, Lockhart DJ, Lerner RA, et al. (2000) Mitotic misregulation and human aging. Science 287: 2486-2492. doi: 10.1126/science.287.5462.2486
|
| [49] | Ferrington DA, Husom AD, Thompson LV (2005) Altered proteasome structure, function, and oxidation in aged muscle. FASEB J 19: 644-646. |
| [50] |
Vernace VA, Arnaud L, Schmidt-Glenewinkel T, et al. (2007) Aging perturbs 26S proteasome assembly in Drosophila melanogaster. FASEB J 21: 2672-2682. doi: 10.1096/fj.06-6751com
|
| [51] |
Grune T, Jung T, Merker K, et al. (2004) Decreased proteolysis caused by protein aggregates, inclusion bodies, plaques, lipofuscin, ceroid, and 'aggresomes' during oxidative stress, aging, and disease. Int J Biochem Cell Biol 36: 2519-2530. doi: 10.1016/j.biocel.2004.04.020
|
| [52] | Andersson V, Hanzen S, Liu B, et al. (2013) Enhancing protein disaggregation restores proteasome activity in aged cells. Aging (Albany NY) 11: 802-812. |
| [53] |
Bulteau AL, Petropoulos I, Friguet B (2000) Age-related alterations of proteasome structure and function in aging epidermis. Exp Gerontol 35: 767-777. doi: 10.1016/S0531-5565(00)00136-4
|
| [54] |
Carrard G, Dieu M, Raes M, et al. (2003) Impact of ageing on proteasome structure and function in human lymphocytes. Int J Biochem Cell Biol 35: 728-739. doi: 10.1016/S1357-2725(02)00356-4
|
| [55] |
Chondrogianni N, Stratford FL, Trougakos IP, et al. (2003) Central role of the proteasome in senescence and survival of human fibroblasts: induction of a senescence-like phenotype upon its inhibition and resistance to stress upon its activation. J Biol Chem 278: 28026-28037. doi: 10.1074/jbc.M301048200
|
| [56] |
Petropoulos I, Conconi M, Wang X, et al. (2000) Increase of oxidatively modified protein is associated with a decrease of proteasome activity and content in aging epidermal cells. J Gerontol A Biol Sci Med Sci 55: B220-227. doi: 10.1093/gerona/55.5.B220
|
| [57] |
Wagner BJ, Margolis JW (1995) Age-dependent association of isolated bovine lens multicatalytic proteinase complex (proteasome) with heat-shock protein 90, an endogenous inhibitor. Arch Biochem Biophys 323: 455-462. doi: 10.1006/abbi.1995.0067
|
| [58] |
Bardag-Gorce F, Farout L, Veyrat-Durebex C, et al. (1999) Changes in 20S proteasome activity during ageing of the LOU rat. Mol Biol Rep 26: 89-93. doi: 10.1023/A:1006968208077
|
| [59] |
Bulteau AL, Szweda LI, Friguet B (2002) Age-dependent declines in proteasome activity in the heart. Arch Biochem Biophys 397: 298-304. doi: 10.1006/abbi.2001.2663
|
| [60] |
Conconi M, Szweda LI, Levine RL, et al. (1996) Age-related decline of rat liver multicatalytic proteinase activity and protection from oxidative inactivation by heat-shock protein 90. Arch Biochem Biophys 331: 232-240. doi: 10.1006/abbi.1996.0303
|
| [61] |
Husom AD, Peters EA, Kolling EA, et al. (2004) Altered proteasome function and subunit composition in aged muscle. Arch Biochem Biophys 421: 67-76. doi: 10.1016/j.abb.2003.10.010
|
| [62] |
Keller JN, Hanni KB, Markesbery WR (2000) Possible involvement of proteasome inhibition in aging: implications for oxidative stress. Mech Ageing Dev 113: 61-70. doi: 10.1016/S0047-6374(99)00101-3
|
| [63] | Shibatani T, Nazir M, Ward WF (1996) Alteration of rat liver 20S proteasome activities by age and food restriction. J Gerontol A Biol Sci Med Sci 51: B316-322. |
| [64] |
Tomaru U, Takahashi S, Ishizu A, et al. (2012) Decreased proteasomal activity causes age-related phenotypes and promotes the development of metabolic abnormalities. Am J Pathol 180: 963-972. doi: 10.1016/j.ajpath.2011.11.012
|
| [65] |
Bergamini E (2006) Autophagy: a cell repair mechanism that retards ageing and age-associated diseases and can be intensified pharmacologically. Mol Aspects Med 27: 403-410. doi: 10.1016/j.mam.2006.08.001
|
| [66] |
Cuervo AM (2008) Autophagy and aging: keeping that old broom working. Trends Genet 24: 604-612. doi: 10.1016/j.tig.2008.10.002
|
| [67] |
Cuervo AM, Dice JF (2000) Age-related decline in chaperone-mediated autophagy. J Biol Chem 275: 31505-31513. doi: 10.1074/jbc.M002102200
|
| [68] | Terman A (1995) The effect of age on formation and elimination of autophagic vacuoles in mouse hepatocytes. Gerontology 41 Suppl 2: 319-326. |
| [69] |
Vittorini S, Paradiso C, Donati A, et al. (1999) The age-related accumulation of protein carbonyl in rat liver correlates with the age-related decline in liver proteolytic activities. J Gerontol A Biol Sci Med Sci 54: B318-323. doi: 10.1093/gerona/54.8.B318
|
| [70] |
Terman A, Brunk UT (2006) Oxidative stress, accumulation of biological 'garbage', and aging. Antioxid Redox Signal 8: 197-204. doi: 10.1089/ars.2006.8.197
|
| [71] |
Yen WL, Klionsky DJ (2008) How to live long and prosper: autophagy, mitochondria, and aging. Physiology (Bethesda) 23: 248-262. doi: 10.1152/physiol.00013.2008
|
| [72] |
Lipinski MM, Zheng B, Lu T, et al. (2010) Genome-wide analysis reveals mechanisms modulating autophagy in normal brain aging and in Alzheimer's disease. Proc Natl Acad Sci U S A 107: 14164-14169. doi: 10.1073/pnas.1009485107
|
| [73] | Carames B, Taniguchi N, Otsuki S, et al. (2010) Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum 62: 791-801. |
| [74] |
Donati A, Cavallini G, Paradiso C, et al. (2001) Age-related changes in the regulation of autophagic proteolysis in rat isolated hepatocytes. J Gerontol A Biol Sci Med Sci 56: B288-293. doi: 10.1093/gerona/56.7.B288
|
| [75] |
Matecic M, Smith DL, Pan X, et al. (2010) A microarray-based genetic screen for yeast chronological aging factors. PLoS Genet 6: e1000921. doi: 10.1371/journal.pgen.1000921
|
| [76] |
Hars ES, Qi H, Ryazanov AG, et al. (2007) Autophagy regulates ageing in C. elegans. Autophagy 3: 93-95. doi: 10.4161/auto.3636
|
| [77] |
Toth ML, Sigmond T, Borsos E, et al. (2008) Longevity pathways converge on autophagy genes to regulate life span in Caenorhabditis elegans. Autophagy 4: 330-338. doi: 10.4161/auto.5618
|
| [78] |
Simonsen A, Cumming RC, Brech A, et al. (2008) Promoting basal levels of autophagy in the nervous system enhances longevity and oxidant resistance in adult Drosophila. Autophagy 4: 176-184. doi: 10.4161/auto.5269
|
| [79] |
Kuma A, Hatano M, Matsui M, et al. (2004) The role of autophagy during the early neonatal starvation period. Nature 432: 1032-1036. doi: 10.1038/nature03029
|
| [80] |
Levine B, Kroemer G (2008) Autophagy in the pathogenesis of disease. Cell 132: 27-42. doi: 10.1016/j.cell.2007.12.018
|
| [81] |
Hara T, Nakamura K, Matsui M, et al. (2006) Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441: 885-889. doi: 10.1038/nature04724
|
| [82] |
Komatsu M, Waguri S, Chiba T, et al. (2006) Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441: 880-884. doi: 10.1038/nature04723
|
| [83] |
Komatsu M, Wang QJ, Holstein GR, et al. (2007) Essential role for autophagy protein Atg7 in the maintenance of axonal homeostasis and the prevention of axonal degeneration. Proc Natl Acad Sci U S A 104: 14489-14494. doi: 10.1073/pnas.0701311104
|
| [84] |
Liang CC, Wang C, Peng X, et al. (2010) Neural-specific deletion of FIP200 leads to cerebellar degeneration caused by increased neuronal death and axon degeneration. J Biol Chem 285: 3499-3509. doi: 10.1074/jbc.M109.072389
|
| [85] |
Masiero E, Agatea L, Mammucari C, et al. (2009) Autophagy is required to maintain muscle mass. Cell Metab 10: 507-515. doi: 10.1016/j.cmet.2009.10.008
|
| [86] |
Singh R, Kaushik S, Wang Y, et al. (2009) Autophagy regulates lipid metabolism. Nature 458: 1131-1135. doi: 10.1038/nature07976
|
| [87] |
Yang L, Li P, Fu S, et al. (2010) Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab 11: 467-478. doi: 10.1016/j.cmet.2010.04.005
|
| [88] |
Jung HS, Chung KW, Won Kim J, et al. (2008) Loss of autophagy diminishes pancreatic beta cell mass and function with resultant hyperglycemia. Cell Metab 8: 318-324. doi: 10.1016/j.cmet.2008.08.013
|
| [89] |
Hartleben B, Godel M, Meyer-Schwesinger C, et al. (2010) Autophagy influences glomerular disease susceptibility and maintains podocyte homeostasis in aging mice. J Clin Invest 120: 1084-1096. doi: 10.1172/JCI39492
|
| [90] | Dice JF (1982) Altered degradation of proteins microinjected into senescent human fibroblasts. J Biol Chem 257: 14624-14627. |
| [91] |
Kiffin R, Kaushik S, Zeng M, et al. (2007) Altered dynamics of the lysosomal receptor for chaperone-mediated autophagy with age. J Cell Sci 120: 782-791. doi: 10.1242/jcs.001073
|
| [92] |
Zhang C, Cuervo AM (2008) Restoration of chaperone-mediated autophagy in aging liver improves cellular maintenance and hepatic function. Nat Med 14: 959-965. doi: 10.1038/nm.1851
|
| [93] |
Bandyopadhyay U, Kaushik S, Varticovski L, et al. (2008) The chaperone-mediated autophagy receptor organizes in dynamic protein complexes at the lysosomal membrane. Mol Cell Biol 28: 5747-5763. doi: 10.1128/MCB.02070-07
|
| [94] |
Soti C, Csermely P (2003) Aging and molecular chaperones. Exp Gerontol 38: 1037-1040. doi: 10.1016/S0531-5565(03)00185-2
|
| [95] |
Nardai G, Csermely P, Soti C (2002) Chaperone function and chaperone overload in the aged. A preliminary analysis. Exp Gerontol 37: 1257-1262. doi: 10.1016/S0531-5565(02)00134-1
|
| [96] |
Cohen E, Dillin A (2008) The insulin paradox: aging, proteotoxicity and neurodegeneration. Nat Rev Neurosci 9: 759-767. doi: 10.1038/nrn2474
|
| [97] |
Zabel C, Nguyen HP, Hin SC, et al. (2010) Proteasome and oxidative phoshorylation changes may explain why aging is a risk factor for neurodegenerative disorders. J Proteomics 73: 2230-2238. doi: 10.1016/j.jprot.2010.08.008
|
| [98] | Matsuda N, Tanaka K (2010) Does impairment of the ubiquitin-proteasome system or the autophagy-lysosome pathway predispose individuals to neurodegenerative disorders such as Parkinson's disease? J Alzheimers Dis 19: 1-9. |
| [99] |
Saez I, Vilchez D (2014) The Mechanistic Links Between Proteasome Activity, Aging and Age-related Diseases. Curr Genomics 15: 38-51. doi: 10.2174/138920291501140306113344
|
| [100] |
Massey AC, Kaushik S, Sovak G, et al. (2006) Consequences of the selective blockage of chaperone-mediated autophagy. Proc Natl Acad Sci U S A 103: 5805-5810. doi: 10.1073/pnas.0507436103
|
| [101] |
Ravikumar B, Vacher C, Berger Z, et al. (2004) Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat Genet 36: 585-595. doi: 10.1038/ng1362
|
| [102] |
Cuervo AM (2006) Autophagy in neurons: it is not all about food. Trends Mol Med 12: 461-464. doi: 10.1016/j.molmed.2006.08.003
|
| [103] |
Keck S, Nitsch R, Grune T, et al. (2003) Proteasome inhibition by paired helical filament-tau in brains of patients with Alzheimer's disease. J Neurochem 85: 115-122. doi: 10.1046/j.1471-4159.2003.01642.x
|
| [104] |
Grune T, Botzen D, Engels M, et al. (2010) Tau protein degradation is catalyzed by the ATP/ubiquitin-independent 20S proteasome under normal cell conditions. Arch Biochem Biophys 500: 181-188. doi: 10.1016/j.abb.2010.05.008
|
| [105] |
Dickey CA, Koren J, Zhang YJ, et al. (2008) Akt and CHIP coregulate tau degradation through coordinated interactions. Proc Natl Acad Sci U S A 105: 3622-3627. doi: 10.1073/pnas.0709180105
|
| [106] |
Petrucelli L, Dickson D, Kehoe K, et al. (2004) CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum Mol Genet 13: 703-714. doi: 10.1093/hmg/ddh083
|
| [107] |
Dange T, Smith D, Noy T, et al. (2011) Blm10 protein promotes proteasomal substrate turnover by an active gating mechanism. J Biol Chem 286: 42830-42839. doi: 10.1074/jbc.M111.300178
|
| [108] |
Jinwal UK, Koren J, 3rd, Borysov SI, et al. (2010) The Hsp90 cochaperone, FKBP51, increases Tau stability and polymerizes microtubules. J Neurosci 30: 591-599. doi: 10.1523/JNEUROSCI.4815-09.2010
|
| [109] |
Blair LJ, Nordhues BA, Hill SE, et al. (2013) Accelerated neurodegeneration through chaperone-mediated oligomerization of tau. J Clin Invest 123: 4158-4169. doi: 10.1172/JCI69003
|
| [110] | Nixon RA, Wegiel J, Kumar A, et al. (2005) Extensive involvement of autophagy in Alzheimer disease: an immuno-electron microscopy study. J Neuropathol Exp Neurol 64: 113-122. |
| [111] | Berger Z, Ravikumar B, Menzies FM, et al. (2006) Rapamycin alleviates toxicity of different aggregate-prone proteins. Hum Mol Genet 15: 433-442. |
| [112] |
Caccamo A, Majumder S, Richardson A, et al. (2010) Molecular interplay between mammalian target of rapamycin (mTOR), amyloid-beta, and Tau: effects on cognitive impairments. J Biol Chem 285: 13107-13120. doi: 10.1074/jbc.M110.100420
|
| [113] |
Tseng BP, Green KN, Chan JL, et al. (2008) Abeta inhibits the proteasome and enhances amyloid and tau accumulation. Neurobiol Aging 29: 1607-1618. doi: 10.1016/j.neurobiolaging.2007.04.014
|
| [114] |
Yu WH, Cuervo AM, Kumar A, et al. (2005) Macroautophagy--a novel Beta-amyloid peptide-generating pathway activated in Alzheimer's disease. J Cell Biol 171: 87-98. doi: 10.1083/jcb.200505082
|
| [115] |
Cataldo AM, Barnett JL, Berman SA, et al. (1995) Gene expression and cellular content of cathepsin D in Alzheimer's disease brain: evidence for early up-regulation of the endosomal-lysosomal system. Neuron 14: 671-680. doi: 10.1016/0896-6273(95)90324-0
|
| [116] |
Boland B, Kumar A, Lee S, et al. (2008) Autophagy induction and autophagosome clearance in neurons: relationship to autophagic pathology in Alzheimer's disease. J Neurosci 28: 6926-6937. doi: 10.1523/JNEUROSCI.0800-08.2008
|
| [117] |
Rozmahel R, Huang J, Chen F, et al. (2002) Normal brain development in PS1 hypomorphic mice with markedly reduced gamma-secretase cleavage of betaAPP. Neurobiol Aging 23: 187-194. doi: 10.1016/S0197-4580(01)00267-6
|
| [118] | Pickford F, Masliah E, Britschgi M, et al. (2008) The autophagy-related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid beta accumulation in mice. J Clin Invest 118: 2190-2199. |
| [119] |
Jaeger PA, Pickford F, Sun CH, et al. (2010) Regulation of amyloid precursor protein processing by the Beclin 1 complex. PLoS One 5: e11102. doi: 10.1371/journal.pone.0011102
|
| [120] |
McNaught KS, Perl DP, Brownell AL, et al. (2004) Systemic exposure to proteasome inhibitors causes a progressive model of Parkinson's disease. Ann Neurol 56: 149-162. doi: 10.1002/ana.20186
|
| [121] |
Bedford L, Hay D, Devoy A, et al. (2008) Depletion of 26S proteasomes in mouse brain neurons causes neurodegeneration and Lewy-like inclusions resembling human pale bodies. J Neurosci 28: 8189-8198. doi: 10.1523/JNEUROSCI.2218-08.2008
|
| [122] |
Wahl C, Kautzmann S, Krebiehl G, et al. (2008) A comprehensive genetic study of the proteasomal subunit S6 ATPase in German Parkinson's disease patients. J Neural Transm 115: 1141-1148. doi: 10.1007/s00702-008-0054-3
|
| [123] |
Tofaris GK, Layfield R, Spillantini MG (2001) alpha-synuclein metabolism and aggregation is linked to ubiquitin-independent degradation by the proteasome. FEBS Lett 509: 22-26. doi: 10.1016/S0014-5793(01)03115-5
|
| [124] | Anglade P, Vyas S, Javoy-Agid F, et al. (1997) Apoptosis and autophagy in nigral neurons of patients with Parkinson's disease. Histol Histopathol 12: 25-31. |
| [125] |
Vogiatzi T, Xilouri M, Vekrellis K, et al. (2008) Wild type alpha-synuclein is degraded by chaperone-mediated autophagy and macroautophagy in neuronal cells. J Biol Chem 283: 23542-23556. doi: 10.1074/jbc.M801992200
|
| [126] |
Webb JL, Ravikumar B, Atkins J, et al. (2003) Alpha-Synuclein is degraded by both autophagy and the proteasome. J Biol Chem 278: 25009-25013. doi: 10.1074/jbc.M300227200
|
| [127] |
Cuervo AM, Stefanis L, Fredenburg R, et al. (2004) Impaired degradation of mutant alpha-synuclein by chaperone-mediated autophagy. Science 305: 1292-1295. doi: 10.1126/science.1101738
|
| [128] |
Spencer B, Potkar R, Trejo M, et al. (2009) Beclin 1 gene transfer activates autophagy and ameliorates the neurodegenerative pathology in alpha-synuclein models of Parkinson's and Lewy body diseases. J Neurosci 29: 13578-13588. doi: 10.1523/JNEUROSCI.4390-09.2009
|
| [129] |
Narendra D, Tanaka A, Suen DF, et al. (2008) Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J Cell Biol 183: 795-803. doi: 10.1083/jcb.200809125
|
| [130] |
Narendra DP, Jin SM, Tanaka A, et al. (2010) PINK1 is selectively stabilized on impaired mitochondria to activate Parkin. PLoS Biol 8: e1000298. doi: 10.1371/journal.pbio.1000298
|
| [131] |
Tashiro Y, Urushitani M, Inoue H, et al. (2012) Motor neuron-specific disruption of proteasomes, but not autophagy, replicates amyotrophic lateral sclerosis. J Biol Chem 287: 42984-42994. doi: 10.1074/jbc.M112.417600
|
| [132] |
Gusella JF, MacDonald ME, Ambrose CM, et al. (1993) Molecular genetics of Huntington's disease. Arch Neurol 50: 1157-1163. doi: 10.1001/archneur.1993.00540110037003
|
| [133] |
Vonsattel JP, DiFiglia M (1998) Huntington disease. J Neuropathol Exp Neurol 57: 369-384. doi: 10.1097/00005072-199805000-00001
|
| [134] | (1993) A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's Disease Collaborative Research Group. Cell 72: 971-983. |
| [135] |
Sanchez I, Mahlke C, Yuan J (2003) Pivotal role of oligomerization in expanded polyglutamine neurodegenerative disorders. Nature 421: 373-379. doi: 10.1038/nature01301
|
| [136] |
Snell RG, MacMillan JC, Cheadle JP, et al. (1993) Relationship between trinucleotide repeat expansion and phenotypic variation in Huntington's disease. Nat Genet 4: 393-397. doi: 10.1038/ng0893-393
|
| [137] |
Nance MA, Myers RH (2001) Juvenile onset Huntington's disease--clinical and research perspectives. Ment Retard Dev Disabil Res Rev 7: 153-157. doi: 10.1002/mrdd.1022
|
| [138] |
Holmberg CI, Staniszewski KE, Mensah KN, et al. (2004) Inefficient degradation of truncated polyglutamine proteins by the proteasome. EMBO J 23: 4307-4318. doi: 10.1038/sj.emboj.7600426
|
| [139] |
Seo H, Sonntag KC, Kim W, et al. (2007) Proteasome activator enhances survival of Huntington's disease neuronal model cells. PLoS One 2: e238. doi: 10.1371/journal.pone.0000238
|
| [140] | Vilchez D, Morantte I, Liu Z, et al. (2012) RPN-6 determines C. elegans longevity under proteotoxic stress conditions. Nature 489: 263-268. |
| [141] |
Tonoki A, Kuranaga E, Tomioka T, et al. (2009) Genetic evidence linking age-dependent attenuation of the 26S proteasome with the aging process. Mol Cell Biol 29: 1095-1106. doi: 10.1128/MCB.01227-08
|
| [142] |
Hipp MS, Patel CN, Bersuker K, et al. (2012) Indirect inhibition of 26S proteasome activity in a cellular model of Huntington's disease. J Cell Biol 196: 573-587. doi: 10.1083/jcb.201110093
|
| [143] |
Kalchman MA, Graham RK, Xia G, et al. (1996) Huntingtin is ubiquitinated and interacts with a specific ubiquitin-conjugating enzyme. J Biol Chem 271: 19385-19394. doi: 10.1074/jbc.271.32.19385
|
| [144] |
Finkbeiner S, Mitra S (2008) The ubiquitin-proteasome pathway in Huntington's disease. ScientificWorldJournal 8: 421-433. doi: 10.1100/tsw.2008.60
|
| [145] |
Sieradzan KA, Mechan AO, Jones L, et al. (1999) Huntington's disease intranuclear inclusions contain truncated, ubiquitinated huntingtin protein. Exp Neurol 156: 92-99. doi: 10.1006/exnr.1998.7005
|
| [146] |
Waelter S, Boeddrich A, Lurz R, et al. (2001) Accumulation of mutant huntingtin fragments in aggresome-like inclusion bodies as a result of insufficient protein degradation. Mol Biol Cell 12: 1393-1407. doi: 10.1091/mbc.12.5.1393
|
| [147] |
Ardley HC, Hung CC, Robinson PA (2005) The aggravating role of the ubiquitin-proteasome system in neurodegeneration. FEBS Lett 579: 571-576. doi: 10.1016/j.febslet.2004.12.058
|
| [148] |
Ciechanover A, Brundin P (2003) The ubiquitin proteasome system in neurodegenerative diseases: sometimes the chicken, sometimes the egg. Neuron 40: 427-446. doi: 10.1016/S0896-6273(03)00606-8
|
| [149] |
Venkatraman P, Wetzel R, Tanaka M, et al. (2004) Eukaryotic proteasomes cannot digest polyglutamine sequences and release them during degradation of polyglutamine-containing proteins. Mol Cell 14: 95-104. doi: 10.1016/S1097-2765(04)00151-0
|
| [150] |
Bennett EJ, Bence NF, Jayakumar R, et al. (2005) Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Mol Cell 17: 351-365. doi: 10.1016/j.molcel.2004.12.021
|
| [151] |
Pratt G, Rechsteiner M (2008) Proteasomes cleave at multiple sites within polyglutamine tracts: activation by PA28gamma(K188E). J Biol Chem 283: 12919-12925. doi: 10.1074/jbc.M709347200
|
| [152] |
Tellez-Nagel I, Johnson AB, Terry RD (1974) Studies on brain biopsies of patients with Huntington's chorea. J Neuropathol Exp Neurol 33: 308-332. doi: 10.1097/00005072-197404000-00008
|
| [153] |
Qin ZH, Wang Y, Kegel KB, et al. (2003) Autophagy regulates the processing of amino terminal huntingtin fragments. Hum Mol Genet 12: 3231-3244. doi: 10.1093/hmg/ddg346
|
| [154] |
Ravikumar B, Duden R, Rubinsztein DC (2002) Aggregate-prone proteins with polyglutamine and polyalanine expansions are degraded by autophagy. Hum Mol Genet 11: 1107-1117. doi: 10.1093/hmg/11.9.1107
|
| [155] |
Sarkar S, Perlstein EO, Imarisio S, et al. (2007) Small molecules enhance autophagy and reduce toxicity in Huntington's disease models. Nat Chem Biol 3: 331-338. doi: 10.1038/nchembio883
|
| [156] |
Martinez-Vicente M, Talloczy Z, Wong E, et al. (2010) Cargo recognition failure is responsible for inefficient autophagy in Huntington's disease. Nat Neurosci 13: 567-576. doi: 10.1038/nn.2528
|
| [157] |
Shibata M, Lu T, Furuya T, et al. (2006) Regulation of intracellular accumulation of mutant Huntingtin by Beclin 1. J Biol Chem 281: 14474-14485. doi: 10.1074/jbc.M600364200
|
| [158] |
Atwal RS, Truant R (2008) A stress sensitive ER membrane-association domain in Huntingtin protein defines a potential role for Huntingtin in the regulation of autophagy. Autophagy 4: 91-93. doi: 10.4161/auto.5201
|
| [159] |
Cattaneo E, Zuccato C, Tartari M (2005) Normal huntingtin function: an alternative approach to Huntington's disease. Nat Rev Neurosci 6: 919-930. doi: 10.1038/nrn1806
|
| [160] |
Ravikumar B, Acevedo-Arozena A, Imarisio S, et al. (2005) Dynein mutations impair autophagic clearance of aggregate-prone proteins. Nat Genet 37: 771-776. doi: 10.1038/ng1591
|
| [161] |
Hassan WM, Merin DA, Fonte V, et al. (2009) AIP-1 ameliorates beta-amyloid peptide toxicity in a Caenorhabditis elegans Alzheimer's disease model. Hum Mol Genet 18: 2739-2747. doi: 10.1093/hmg/ddp209
|
| [162] |
Yun C, Stanhill A, Yang Y, et al. (2008) Proteasomal adaptation to environmental stress links resistance to proteotoxicity with longevity in Caenorhabditis elegans. Proc Natl Acad Sci U S A 105: 7094-7099. doi: 10.1073/pnas.0707025105
|
| [163] |
Nakatogawa H, Suzuki K, Kamada Y, et al. (2009) Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nat Rev Mol Cell Biol 10: 458-467. doi: 10.1038/nrm2708
|
| [164] |
Xie Z, Klionsky DJ (2007) Autophagosome formation: core machinery and adaptations. Nat Cell Biol 9: 1102-1109. doi: 10.1038/ncb1007-1102
|
| [165] | Yang Z, Klionsky DJ (2009) An overview of the molecular mechanism of autophagy. Curr Top Microbiol Immunol 335: 1-32. |
| [166] |
Hanada T, Noda NN, Satomi Y, et al. (2007) The Atg12-Atg5 conjugate has a novel E3-like activity for protein lipidation in autophagy. J Biol Chem 282: 37298-37302. doi: 10.1074/jbc.C700195200
|
| [167] |
Hanada T, Satomi Y, Takao T, et al. (2009) The amino-terminal region of Atg3 is essential for association with phosphatidylethanolamine in Atg8 lipidation. FEBS Lett 583: 1078-1083. doi: 10.1016/j.febslet.2009.03.009
|
| [168] |
Komatsu M, Waguri S, Ueno T, et al. (2005) Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J Cell Biol 169: 425-434. doi: 10.1083/jcb.200412022
|
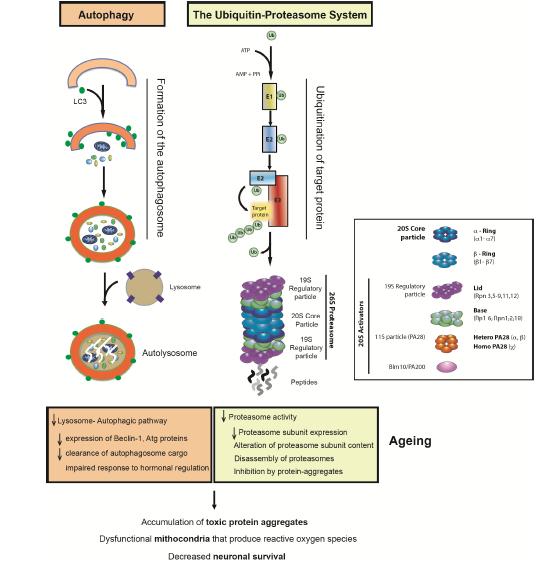
Figures(1) / Tables(1)

Isabel Saez, David Vilchez. Protein clearance mechanisms and their demise in age-related neurodegenerative diseases[J]. AIMS Molecular Science, 2015, 1(1): 1-21. doi: 10.3934/molsci.2015.1.1







 DownLoad:
DownLoad: