The human oral microbiome can affect brain functions directly through the trigeminal nerve and olfactory system and indirectly via the oral–gut–brain axis. However, the potential link between the oral microbiome and mental health remains an area that requires further investigation. Taking into consideration that gut microbiota dysbiosis plays a role in the onset and progression of several mental disorders, as well as the potential influence of the oral microbiome on mental health via direct pathways, the present narrative review explores the role of the human oral microbiome in health and disease, along with the factors that affect its composition, with a particular focus on its potential impact on mental health, including its involvement in a range of mental disorders and brain-related conditions, such as Alzheimer's disease, Parkinson's disease, autism spectrum disorder, anxiety, depression, stress, bipolar disorder, Down's syndrome, cerebral palsy, epilepsy, and schizophrenia. Chronic oral diseases can impair the oral mucosal barrier, allowing microorganisms and endotoxins to enter the bloodstream, triggering systemic inflammation, and affecting the blood–brain barrier. This pathway can lead to neuroinflammation and cognitive dysfunction and contribute to adverse mental health effects. Additionally, translocation of oral bacteria to the gut can drive persistent inflammation and thereby affect brain health. Multiple studies suggest a potential relationship between the oral microbiome and several mental disorders, but further research is needed to strengthen the evidence surrounding these associations and to fully clarify the underlying mechanisms linking the oral microbiome to these conditions. Given the promising implications, future research should focus on elucidating the biological mechanisms through which alterations in the oral microbiome influence the development and progression of determinate neurodegenerative and neuropsychiatric disorders. Additionally, identifying reliable biomarkers linked to the oral microbiome could enhance early detection, diagnosis, and monitoring of these conditions.
Citation: Alejandro Borrego-Ruiz, Juan J. Borrego. Human oral microbiome and its influence on mental health and brain disorders[J]. AIMS Microbiology, 2025, 11(2): 242-294. doi: 10.3934/microbiol.2025013
The human oral microbiome can affect brain functions directly through the trigeminal nerve and olfactory system and indirectly via the oral–gut–brain axis. However, the potential link between the oral microbiome and mental health remains an area that requires further investigation. Taking into consideration that gut microbiota dysbiosis plays a role in the onset and progression of several mental disorders, as well as the potential influence of the oral microbiome on mental health via direct pathways, the present narrative review explores the role of the human oral microbiome in health and disease, along with the factors that affect its composition, with a particular focus on its potential impact on mental health, including its involvement in a range of mental disorders and brain-related conditions, such as Alzheimer's disease, Parkinson's disease, autism spectrum disorder, anxiety, depression, stress, bipolar disorder, Down's syndrome, cerebral palsy, epilepsy, and schizophrenia. Chronic oral diseases can impair the oral mucosal barrier, allowing microorganisms and endotoxins to enter the bloodstream, triggering systemic inflammation, and affecting the blood–brain barrier. This pathway can lead to neuroinflammation and cognitive dysfunction and contribute to adverse mental health effects. Additionally, translocation of oral bacteria to the gut can drive persistent inflammation and thereby affect brain health. Multiple studies suggest a potential relationship between the oral microbiome and several mental disorders, but further research is needed to strengthen the evidence surrounding these associations and to fully clarify the underlying mechanisms linking the oral microbiome to these conditions. Given the promising implications, future research should focus on elucidating the biological mechanisms through which alterations in the oral microbiome influence the development and progression of determinate neurodegenerative and neuropsychiatric disorders. Additionally, identifying reliable biomarkers linked to the oral microbiome could enhance early detection, diagnosis, and monitoring of these conditions.

| [1] |
Mark Welch JL, Ramírez-Puebla ST, Borisy GG (2020) Oral microbiome geography: Micron-scale habitat and niche. Cell Host Microbe 28: 160-168. https://doi.org/10.1016/j.chom.2020.07.009 
|
| [2] |
Cho YD, Kim KH, Lee YM, et al. (2021) Oral microbiome and host health: Review on current advances in genome-wide analysis. Appl Sci 11: 4050. https://doi.org/10.3390/app11094050
|
| [3] |
Chowdhry A, Kapoor P, Bhargava D, et al. (2023) Exploring the oral microbiome: An updated multidisciplinary oral healthcare perspective. Discoveries 11: e165. https://doi.org/10.15190/d.2023.4
|
| [4] |
Gao L, Xu T, Huang G, et al. (2018) Oral microbiomes: More and more importance in oral cavity and whole body. Protein Cell 9: 488-500. https://doi.org/10.1007/s13238-018-0548-1
|
| [5] |
Spatafora G, Li Y, He X, et al. (2024) The evolving microbiome of dental caries. Microorganisms 12: 121. https://doi.org/10.3390/microorganisms12010121
|
| [6] |
Cornejo Ulloa P, van der Veen MH, Krom BP (2019) Review: Modulation of the oral microbiome by the host to promote ecological balance. Odontology 107: 437-448. https://doi.org/10.1007/s10266-019-00413-x
|
| [7] |
Bacali C, Vulturar R, Buduru S, et al. (2022) Oral microbiome: Getting to know and befriend neighbors, a biological approach. Biomedicines 10: 671. https://doi.org/10.3390/biomedicines10030671
|
| [8] |
Dethlefsen L, McFall-Ngai M, Relman DA (2007) An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 449: 811-818. https://doi.org/10.1038/nature06245
|
| [9] |
Bäumler AJ, Sperandio V (2016) Interactions between the microbiota and pathogenic bacteria in the gut. Nature 535: 85-93. https://doi.org/10.1038/nature18849
|
| [10] |
Honda K, Littman DR (2016) The microbiota in adaptive immune homeostasis and disease. Nature 535: 75-84. https://doi.org/10.1038/nature18848
|
| [11] |
Thaiss CA, Zmora N, Levy M, et al. (2016) The microbiome and innate immunity. Nature 535: 65-74. https://doi.org/10.1038/nature18847
|
| [12] |
Kilian M (2018) The oral microbiome-friend or foe?. Eur J Oral Sci 126: 5-12. https://doi.org/10.1111/eos.12527
|
| [13] | Integrative HMP (iHMP) Research Network Consortium.The integrative human microbiome project. Nature (2019) 569: 641-648. https://doi.org/10.1038/s41586-019-1238-8 |
| [14] |
Escapa IF, Chen T, Huang Y, et al. (2018) New insights into human nostril microbiome from the expanded Human Oral Microbiome Database (eHOMD): A resource for the microbiome of the human aerodigestive tract. mSystems 3: e00187-18. https://doi.org/10.1128/mSystems.00187-18
|
| [15] |
Verma D, Garg PK, Dubey AK (2018) Insights into the human oral microbiome. Arch Microbiol 200: 525-540. https://doi.org/10.1007/s00203-018-1505-3
|
| [16] |
Shi H, Shi Q, Grodner B, et al. (2020) Highly multiplexed spatial mapping of microbial communities. Nature 588: 676-681. https://doi.org/10.1038/s41586-020-2983-4
|
| [17] |
Van't Hof W, Veerman EC, Nieuw Amerongen AV, et al. (2014) Antimicrobial defense systems in saliva. Monogr Oral Sci 24: 40-51. https://doi.org/10.1159/000358783
|
| [18] |
Sultan AS, Kong EF, Rizk AM, et al. (2018) The oral microbiome: A lesson in coexistence. PLoS Pathog 14: e1006719. https://doi.org/10.1371/journal.ppat.1006719
|
| [19] |
Camanocha A, Dewhirst FE (2014) Host-associated bacterial taxa from Chlorobi, Chloroflexi, GN02, Synergistetes, SR1, TM7, and WPS-2 Phyla/candidate divisions. J Oral Microbiol 6: 10.3402/jom.v6.25468. https://doi.org/10.3402/jom.v6.25468
|
| [20] |
Campbell JH, O'Donoghue P, Campbell AG, et al. (2013) UGA is an additional glycine codon in uncultured SR1 bacteria from the human microbiota. Proc Natl Acad Sci U S A 110: 5540-5545. https://doi.org/10.1073/pnas.1303090110
|
| [21] |
Fernandes CdC, Rechenberg DK, Zehnder M, et al. (2014) Identification of Synergistetes in endodontic infections. Microb Pathog 73: 1-6. https://doi.org/10.1016/j.micpath.2014.05.001
|
| [22] |
Murugkar PP, Collins AJ, Chen T, et al. (2020) Isolation and cultivation of candidate phyla radiation Saccharibacteria (TM7) bacteria in coculture with bacterial hosts. J Oral Microbiol 12: 1814666. https://doi.org/10.1080/20002297.2020.1814666
|
| [23] |
Aleksandrowicz P, Brzezińska-Błaszczyk E, Dudko A, et al. (2020) Archaea occurrence in the subgingival biofilm in patients with peri-implantitis and periodontitis. Int J Periodontics Restorative Dent 40: 677-683. https://doi.org/10.11607/prd.4670
|
| [24] |
Dame-Teixeira N, de Cena JA, Côrtes DA, et al. (2020) Presence of Archaea in dental caries biofilms. Arch Oral Biol 110: 104606. https://doi.org/10.1016/j.archoralbio.2019.104606
|
| [25] | Santacroce L, Sardaro N, Topi S, et al. (2020) The pivotal role of oral microbiota in health and disease. J Biol Regul Homeost Agents 34: 733-737. https://doi.org/10.23812/20-127-L-45 |
| [26] |
Hong BY, Hoare A, Cardenas A, et al. (2020) The salivary mycobiome contains 2 ecologically distinct mycotypes. J Dent Res 99: 730-738. https://doi.org/10.1177/0022034520915879
|
| [27] |
Peters BA, Wu J, Hayes RB, et al. (2017) The oral fungal mycobiome: Characteristics and relation to periodontitis in a pilot study. BMC Microbiol 17: 157. https://doi.org/10.1186/s12866-017-1064-9
|
| [28] |
Caselli E, Fabbri C, D'Accolti M, et al. (2020) Defining the oral microbiome by whole-genome sequencing and resistome analysis: The complexity of the healthy picture. BMC Microbiol 20: 120. https://doi.org/10.1186/s12866-020-01801-y
|
| [29] |
Morán-Torres A, Pazos-Salazar NG, Téllez-Lorenzo S, et al. (2021) HPV oral and oropharynx infection dynamics in young population. Braz J Microbiol 52: 1991-2000. https://doi.org/10.1007/s42770-021-00602-3
|
| [30] |
Syrjanen S (2018) Oral manifestations of human papillomavirus infections. Eur J Oral Sci 126: 49-66. https://doi.org/10.1111/eos.12538
|
| [31] |
Atyeo N, Rodriguez MD, Papp B, et al. (2021) Clinical manifestations and epigenetic regulation of oral herpesvirus infections. Viruses 13: 681. https://doi.org/10.3390/v13040681
|
| [32] |
Crimi S, Fiorillo L, Bianchi A, et al. (2019) Herpes virus, oral clinical signs and QoL: Systematic review of recent data. Viruses 11: 463. https://doi.org/10.3390/v11050463
|
| [33] |
Polvora TLS, Nobre AVV, Tirapelli C, et al. (2018) Relationship between human immunodeficiency virus (HIV-1) infection and chronic periodontitis. Expert Rev Clin Immunol 14: 315-327. https://doi.org/10.1080/1744666X.2018.1459571
|
| [34] |
Xiao X, Liu S, Deng H, et al. (2023) Advances in the oral microbiota and rapid detection of oral infectious diseases. Front Microbiol 14: 1121737. https://doi.org/10.3389/fmicb.2023.1121737
|
| [35] |
Banar M, Rokaya D, Azizian R, et al. (2024) Oral bacteriophages: Metagenomic clues to interpret microbiomes. Peer J 12: e16947. https://doi.org/10.7717/peerj.16947
|
| [36] |
Edlund A, Santiago-Rodriguez TM, Boehm TK, et al. (2015) Bacteriophage and their potential roles in the human oral cavity. J Oral Microbiol 7: 27423. https://doi.org/10.3402/jom.v7.27423
|
| [37] |
Szafrański SP, Slots J, Stiesch M (2021) The human oral phageome. Periodontol. 2000 86: 79-96. https://doi.org/10.1111/prd.12363
|
| [38] |
Pride DT, Salzman J, Haynes M, et al. (2012) Evidence of a robust resident bacteriophage population revealed through analysis of the human salivary virome. ISME J 6: 915-926. https://doi.org/10.1038/ismej.2011.169
|
| [39] |
Yaseen A, Mahafzah A, Dababseh D, et al. (2021) Oral colonization by Entamoeba gingivalis and Trichomonas tenax: A PCR-based study in health, gingivitis, and periodontitis. Front Cell Infect Microbiol 11: 782805. https://doi.org/10.3389/fcimb.2021.782805
|
| [40] |
Dewhirst FE, Chen T, Izard J, et al. (2010) The human oral microbiome. J Bacteriol 192: 5002-5017. https://doi.org/10.1128/JB.00542-10
|
| [41] |
Seidel CL, Gerlach RG, Wiedemann P, et al. (2020) Defining metaniches in the oral cavity according to their microbial composition and cytokine profile. Int J Mol Sci 21: 8218. https://doi.org/10.3390/ijms21218218
|
| [42] |
Asakawa M, Takeshita T, Furuta M, et al. (2018) Tongue microbiota and oral health status in community-dwelling elderly adults. mSphere 3: e00332-18. https://doi.org/10.1128/mSphere.00332-18
|
| [43] |
Camelo-Castillo AJ, Mira A, Pico A, et al. (2015) Subgingival microbiota in health compared to periodontitis and the influence of smoking. Front Microbiol 6: 119. https://doi.org/10.3389/fmicb.2015.00119
|
| [44] |
Espinoza JL, Harkins DM, Torralba M, et al. (2018) Supragingival plaque microbiome ecology and functional potential in the context of health and disease. mBio 9: e01631-18. https://doi.org/10.1128/mBio.01631-18
|
| [45] |
Inquimbert C, Bourgeois D, Bravo M, et al. (2019) The oral bacterial microbiome of interdental surfaces in adolescents according to carious risk. Microorganisms 7: 319. https://doi.org/10.3390/microorganisms7090319
|
| [46] |
Lee YH, Chung SW, Auh QS, et al. (2021) Progress in oral microbiome related to oral and systemic diseases: An update. Diagnostics 11: 1283. https://doi.org/10.3390/diagnostics11071283
|
| [47] |
Li X, Liu Y, Yang X, et al. (2022) The oral microbiota: Community composition, influencing factors, pathogenesis, and interventions. Front Microbiol 13: 895537. https://doi.org/10.3389/fmicb.2022.895537
|
| [48] |
Zhang Y, Wang X, Li H, et al. (2018) Human oral microbiota and its modulation for oral health. Biomed Pharmacother 99: 883-893. https://doi.org/10.1016/j.biopha.2018.01.146
|
| [49] |
Shi W, Tian J, Xu H, et al. (2018) Distinctions and associations between the microbiota of saliva and supragingival plaque of permanent and deciduous teeth. PLoS One 13: e0200337. https://doi.org/10.1371/journal.pone.0200337
|
| [50] |
Murugesan S, Al Ahmad SF, Singh P, et al. (2020) Profiling the salivary microbiome of the Qatari population. J Transl Med 18: 127. https://doi.org/10.1186/s12967-020-02291-2
|
| [51] |
Takeshita T, Kageyama S, Furuta M, et al. (2016) Bacterial diversity in saliva and oral health-related conditions: The Hisayama study. Sci Rep 6: 22164. https://doi.org/10.1038/srep22164
|
| [52] |
Shaw L, Ribeiro ALR, Levine AP, et al. (2017) The human salivary microbiome is shaped by shared environment rather than genetics: Evidence from a large family of closely related individuals. mBio 8: e01237-17. https://doi.org/10.1128/mBio.01237-17
|
| [53] |
Lynge Pedersen AM, Belstrøm D (2019) The role of natural salivary defences in maintaining a healthy oral microbiota. J Dent 80: S3-S12. https://doi.org/10.1016/j.jdent.2018.08.010
|
| [54] |
Fábián TK, Hermann P, Beck A, et al. (2012) Salivary defense proteins: Their network and role in innate and acquired oral immunity. Int J Mol Sci 13: 4295-4320. https://doi.org/10.3390/ijms13044295
|
| [55] |
Ren W, Zhang Q, Liu X, et al. (2017) Exploring the oral microflora of preschool children. J Microbiol 55: 531-537. https://doi.org/10.1007/s12275-017-6474-8
|
| [56] |
Costalonga M, Herzberg MC (2014) The oral microbiome and the immunobiology of periodontal disease and caries. Immunol Lett 162: 22-38. https://doi.org/10.1016/j.imlet.2014.08.017
|
| [57] |
Simón-Soro A, Tomás I, Cabrera-Rubio R, et al. (2013) Microbial geography of the oral cavity. J Dent Res 92: 616-621. https://doi.org/10.1177/0022034513488119
|
| [58] |
Takeshita T, Yasui M, Shibata Y, et al. (2015) Dental plaque development on a hydroxyapatite disk in young adults observed by using a barcoded pyrosequencing approach. Sci Rep 5: 8136. https://doi.org/10.1038/srep08136
|
| [59] |
Roberts AP, Kreth J (2014) The impact of horizontal gene transfer on the adaptive ability of the human oral microbiome. Front Cell Infect Microbiol 4: 124. https://doi.org/10.3389/fcimb.2014.00124
|
| [60] |
Kolenbrander PE, Palmer RJ, Periasamy S, et al. (2010) Oral multispecies biofilm development and the key role of cell-cell distance. Nat Rev Microbiol 8: 471-480. https://doi.org/10.1038/nrmicro2381
|
| [61] |
Chen Y, Li Y, Zou J (2019) Intrageneric and intergeneric interactions developed by oral streptococci: Pivotal role in the pathogenesis of oral diseases. Curr Issues Mol Biol 32: 377-434. https://doi.org/10.21775/cimb.032.377
|
| [62] |
Bowen WH, Burne RA, Wu H, et al. (2018) Oral biofilms: Pathogens, matrix, and polymicrobial interactions in microenvironments. Trends Microbiol 26: 229-242. https://doi.org/10.1016/j.tim.2017.09.008
|
| [63] |
Heller D, Helmerhorst EJ, Oppenheim FG (2017) Saliva and serum protein exchange at the tooth enamel surface. J Dent Res 96: 437-443. https://doi.org/10.1177/0022034516680771
|
| [64] |
Mohammed WK, Krasnogor N, Jakubovics NS (2018) Streptococcus gordonii challisin protease is required for sensing cell-cell contact with Actinomyces oris. FEMS Microbiol Ecol 94: fiy043. https://doi.org/10.1093/femsec/fiy043
|
| [65] |
Mutha NVR, Mohammed WK, Krasnogor N, et al. (2018) Transcriptional responses of Streptococcus gordonii and Fusobacterium nucleatum to coaggregation. Mol Oral Microbiol 33: 450-464. https://doi.org/10.1111/omi.12248
|
| [66] |
Takahashi N (2015) Oral microbiome metabolism: From "Who are they?" to "What are they doing?". J Dent Res 94: 1628-1637. https://doi.org/10.1177/0022034515606045
|
| [67] |
Zhou P, Manoil D, Belibasakis GN, et al. (2021) Veillonellae: Beyond bridging species in oral biofilm ecology. Front Oral Health 2: 774115. https://doi.org/10.3389/froh.2021.774115
|
| [68] |
Kriebel K, Hieke C, Muller-Hilke B, et al. (2018) Oral biofilms from symbiotic to pathogenic interactions and associated disease-Connection of periodontitis and rheumatic arthritis by peptidylarginine deiminase. Front Microbiol 9: 53. https://doi.org/10.3389/fmicb.2018.00053
|
| [69] |
Brennan CA, Garrett WS (2019) Fusobacterium nucleatum-Symbiont, opportunist and oncobacterium. Nat Rev Microbiol 17: 156-166. https://doi.org/10.1038/s41579-018-0129-6
|
| [70] |
Lima BP, Shi W, Lux R (2017) Identification and characterization of a novel Fusobacterium nucleatum adhesin involved in physical interaction and biofilm formation with Streptococcus gordonii. Microbiologyopen 6: e00444. https://doi.org/10.1002/mbo3.444
|
| [71] |
Lima BP, Hu LI, Vreeman GW, et al. (2019) The oral bacterium Fusobacterium nucleatum binds Staphylococcus aureus and alters expression of the staphylococcal accessory regulator sarA. Microb Ecol 78: 336-347. https://doi.org/10.1007/s00248-018-1291-0
|
| [72] |
Jayaraman A, Wood TK (2008) Bacterial quorum sensing: Signals, circuits, and implications for biofilms and disease. Annu Rev Biomed Eng 10: 145-167. https://doi.org/10.1146/annurev.bioeng.10.061807.160536
|
| [73] |
Li YH, Tian X (2012) Quorum sensing and bacterial social interactions in biofilms. Sensors 12: 2519-2538. https://doi.org/10.3390/s120302519
|
| [74] |
Santacroce L, Passarelli PC, Azzolino D, et al. (2023) Oral microbiota in human health and disease: A perspective. Exp Biol Med 248: 1288-1301. https://doi.org/10.1177/15353702231187645
|
| [75] |
Belstrøm D, Holmstrup P, Nielsen CH, et al. (2014) Bacterial profiles of saliva in relation to diet, lifestyle factors, and socioeconomic status. J Oral Microbiol 6: 10.3402/jom.v6.23609. https://doi.org/10.3402/jom.v6.23609
|
| [76] |
Dagli N, Dagli R, Darwish S, et al. (2016) Oral microbial shift: Factors affecting the microbiome and prevention of oral disease. J Contemp Dent Pract 17: 90-96. https://doi.org/10.5005/jp-journals-10024-1808
|
| [77] |
Menon RK, Gomez A, Brandt BW, et al. (2019) Long-term impact of oral surgery with or without amoxicillin on the oral microbiome-A prospective cohort study. Sci Rep 9: 18761. https://doi.org/10.1038/s41598-019-55056-3
|
| [78] |
Larsson Wexell C, Ryberg H, Sjöberg Andersson WA, et al. (2016) Antimicrobial effect of a single dose of amoxicillin on the oral microbiota. Clin Implant Dent Relat Res 18: 699-706. https://doi.org/10.1111/cid.12357
|
| [79] |
Moraes LC, Lang PM, Arcanjo RA, et al. (2020) Microbial ecology and predicted metabolic pathways in various oral environments from patients with acute endodontic infections. Int Endod J 53: 1603-1617. https://doi.org/10.1111/iej.13389
|
| [80] |
Raju SC, Viljakainen H, Figueiredo RAO, et al. (2020) Antimicrobial drug use in the first decade of life influences saliva microbiota diversity and composition. Microbiome 8: 121. https://doi.org/10.1186/s40168-020-00893-y
|
| [81] |
Almeida VSM, Azevedo J, Leal HF, et al. (2020) Bacterial diversity and prevalence of antibiotic resistance genes in the oral microbiome. PLoS One 15: e0239664. https://doi.org/10.1371/journal.pone.0239664
|
| [82] |
Oba PM, Holscher HD, Mathai RA, et al. (2020) Diet influences the oral microbiota of infants during the first six months of life. Nutrients 12: 3400. https://doi.org/10.3390/nu12113400
|
| [83] |
Chumponsuk T, Gruneck L, Gentekaki E, et al. (2021) The salivary microbiota of Thai adults with metabolic disorders and association with diet. Arch Oral Biol 122: 105036. https://doi.org/10.1016/j.archoralbio.2020.105036
|
| [84] |
Yussof A, Yoon P, Krkljes C, et al. (2020) A meta-analysis of the effect of binge drinking on the oral microbiome and its relation to Alzheimer's disease. Sci Rep 10: 19872. https://doi.org/10.1038/s41598-020-76784-x
|
| [85] |
Nociti FH, Casati MZ, Duarte PM (2015) Current perspective of the impact of smoking on the progression and treatment of periodontitis. Periodontol 2000 67: 187-210. https://doi.org/10.1111/prd.12063
|
| [86] |
Wu J, Peters BA, Dominianni C, et al. (2016) Cigarette smoking and the oral microbiome in a large study of American adults. ISME J 10: 2435-2446. https://doi.org/10.1038/ismej.2016.37
|
| [87] |
Senaratne NLM, Yung On C, Shetty NY, et al. (2024) Effect of different forms of tobacco on the oral microbiome in healthy adults: A systematic review. Front Oral Health 5: 1310334. https://doi.org/10.3389/froh.2024.1310334
|
| [88] |
Lin W, Jiang W, Hu X, et al. (2018) Ecological shifts of supragingival microbiota in association with pregnancy. Front Cell Infect Microbiol 8: 24. https://doi.org/10.3389/fcimb.2018.00024
|
| [89] |
Fujiwara N, Tsuruda K, Iwamoto Y, et al. (2017) Significant increase of oral bacteria in the early pregnancy period in Japanese women. J Investig Clin Dent 8: e12189. https://doi.org/10.1111/jicd.12189
|
| [90] |
Li H, Xiao B, Zhang Y, et al. (2019) Impact of maternal intrapartum antibiotics on the initial oral microbiome of neonates. Pediatr Neonatol 60: 654-661. https://doi.org/10.1016/j.pedneo.2019.03.011
|
| [91] |
Gao J, Wang H, Li Z, et al. (2018) Candida albicans gains azole resistance by altering sphingolipid composition. Nat Commun 9: 4495. https://doi.org/10.1038/s41467-018-06944-1
|
| [92] |
Monroy-Pérez E, Rodríguez-Bedolla RM, Garzón J, et al. (2020) Marked virulence and azole resistance in Candida albicans isolated from patients with periodontal disease. Microb Pathog 148: 104436. https://doi.org/10.1016/j.micpath.2020.104436
|
| [93] |
Sweeney EL, Al-Shehri SS, Cowley DM, et al. (2018) The effect of breastmilk and saliva combinations on the in vitro growth of oral pathogenic and commensal microorganisms. Sci Rep 8: 15112. https://doi.org/10.1038/s41598-018-33519-3
|
| [94] |
Anderson AC, Rothballer M, Altenburger MJ, et al. (2020) Long-term fluctuation of oral biofilm microbiota following different dietary phases. Appl Environ Microbiol 86: e0142120. https://doi.org/10.1128/AEM.01421-20
|
| [95] |
Hansen TH, Kern T, Bak EG, et al. (2018) Impact of a vegan diet on the human salivary microbiota. Sci Rep 8: 5847. https://doi.org/10.1038/s41598-018-24207-3
|
| [96] |
Štšepetova J, Truu J, Runnel R, et al. (2019) Impact of polyols on oral microbiome of Estonian schoolchildren. BMC Oral Health 19: 60. https://doi.org/10.1186/s12903-019-0747-z
|
| [97] |
Sanchez MC, Ribeiro-Vidal H, Esteban-Fernandez A, et al. (2019) Antimicrobial activity of red wine and oenological extracts against periodontal pathogens in a validated oral biofilm model. BMC Complement Altern Med 19: 145. https://doi.org/10.1186/s12906-019-2533-5
|
| [98] |
Jia YJ, Liao Y, He YQ, et al. (2021) Association between oral microbiota and cigarette smoking in the Chinese population. Front Cell Infect. Microbiol 11: 658203. https://doi.org/10.3389/fcimb.2021.658203
|
| [99] |
Yu G, Phillips S, Gail MH, et al. (2017) The effect of cigarette smoking on the oral and nasal microbiota. Microbiome 5: 3. https://doi.org/10.1186/s40168-016-0226-6
|
| [100] |
Kumar PS, Matthews CR, Joshi V, et al. (2011) Tobacco smoking affects bacterial acquisition and colonization in oral biofilms. Infect Immun 79: 4730-4738. https://doi.org/10.1128/IAI.05371-11
|
| [101] |
Al Bataineh MT, Dash NR, Elkhazendar M, et al. (2020) Revealing oral microbiota composition and functionality associated with heavy cigarette smoking. J Transl Med 18: 421. https://doi.org/10.1186/s12967-020-02579-3
|
| [102] |
Yang Y, Zheng W, Cai QY, et al. (2019) Cigarette smoking and oral microbiota in low-income and African-American populations. J Epidemiol Community Health 73: 1108-1115. https://doi.org/10.1136/jech-2019-212474
|
| [103] |
Saadaoui M, Singh P, Al Khodor S (2021) Oral microbiome and pregnancy: A bidirectional relationship. J Reprod Immunol 145: 103293. https://doi.org/10.1016/j.jri.2021.103293
|
| [104] |
Barak S, Oettinger-Barak O, Oettinger M, et al. (2003) Common oral manifestations during pregnancy: A review. Obstet Gynecol Surv 58: 624-628. https://doi.org/10.1097/01.OGX.0000083542.14439.CF
|
| [105] |
Tettamanti L, Lauritano D, Nardone M, et al. (2017) Pregnancy and periodontal disease: Does exist a two-way relationship?. Oral Implantol 10: 112-118. https://doi.org/10.11138/orl/2017.10.2.112
|
| [106] |
León R, Silva N, Ovalle A, et al. (2007) Detection of Porphyromonas gingivalis in the amniotic fluid in pregnant women with a diagnosis of threatened premature labor. J Periodontol 78: 1249-1255. https://doi.org/10.1902/jop.2007.060368
|
| [107] |
Jang H, Patoine A, Wu TT, et al. (2021) Oral microflora and pregnancy: A systematic review and meta-analysis. Sci Rep 11: 16870. https://doi.org/10.1038/s41598-021-96495-1
|
| [108] |
Pecci-Lloret MP, Linares-Pérez C, Pecci-Lloret MR, et al. (2024) Oral manifestations in pregnant women: A systematic review. J Clin Med 13: 707. https://doi.org/10.3390/jcm13030707
|
| [109] |
Hurley E, Barrett MPJ, Kinirons M, et al. (2019) Comparison of the salivary and dentinal microbiome of children with severe-early childhood caries to the salivary microbiome of caries-free children. BMC Oral Health 19: 13. https://doi.org/10.1186/s12903-018-0693-1
|
| [110] |
Wei Y, Shi M, Zhen M, et al. (2019) Comparison of subgingival and buccal mucosa microbiome in chronic and aggressive periodontitis: A pilot study. Front Cell Infect Microbiol 9: 53. https://doi.org/10.3389/fcimb.2019.00053
|
| [111] |
Kim YT, Jeong J, Mun S, et al. (2022) Comparison of the oral microbial composition between healthy individuals and periodontitis patients in different oral sampling sites using 16S metagenome profiling. J Periodontal Implant Sci 52: 394-410. https://doi.org/10.5051/jpis.2200680034
|
| [112] | Dipalma G, Inchingolo AD, Inchingolo F, et al. (2021) Focus on the cariogenic process: Microbial and biochemical interactions with teeth and oral environment. J Biol Regul Homeost Agents 35: 429-440. https://doi.org/10.23812/20-747-A |
| [113] |
Belibasakis GN, Bostanci N, Marsh PD, et al. (2019) Applications of the oral microbiome in personalized dentistry. Arch Oral Biol 104: 7-12. https://doi.org/10.1016/j.archoralbio.2019.05.023
|
| [114] |
Bek-Thomsen M, Tettelin H, Hance I, et al. (2008) Population diversity and dynamics of Streptococcus mitis, Streptococcus oralis, and Streptococcus infantis in the upper respiratory tracts of adults, determined by a nonculture strategy. Infect Immun 76: 1889-1896. https://doi.org/10.1128/IAI.01511-07
|
| [115] |
López-López A, Camelo-Castillo A, Ferrer MD, et al. (2017) Health-associated niche inhabitants as oral probiotics: The case of Streptococcus dentisani. Front Microbiol 8: 379. https://doi.org/10.3389/fmicb.2017.00379
|
| [116] |
Morou-Bermudez E, Rodriguez S, Bello AS, et al. (2015) Urease and dental plaque microbial profiles in children. PloS One 10: e0139315. https://doi.org/10.1371/journal.pone.0139315
|
| [117] |
Reyes E, Martin J, Moncada G, et al. (2014) Caries-free subjects have high levels of urease and arginine deiminase activity. J Appl Oral Sci 22: 235-240. https://doi.org/10.1590/1678-775720130591
|
| [118] | Tanner AC, Kressirer CA, Faller LL (2016) Understanding caries from the oral microbiome perspective. J Calif Dent Assoc 44: 437-446. |
| [119] |
Ma C, Chen F, Zhang Y, et al. (2015) Comparison of oral microbial profiles between children with severe early childhood caries and caries-free children using the human oral microbe identification microarray. PLoS One 10: e0122075. https://doi.org/10.1371/journal.pone.0122075
|
| [120] |
Wang Y, Zhang J, Chen X, et al. (2017) Profiling of oral microbiota in early childhood caries using single-molecule real-time sequencing. Front Microbiol 8: 2244. https://doi.org/10.3389/fmicb.2017.02244
|
| [121] |
Agnello M, Marques J, Cen L, et al. (2017) Microbiome associated with severe caries in Canadian First Nations children. J Dent Res 96: 1378-1385. https://doi.org/10.1177/22034517718819
|
| [122] |
Giordano-Kelhoffer B, Lorca C, March Llanes J, et al. (2022) Oral microbiota, its equilibrium and implications in the pathophysiology of human diseases: A systematic review. Biomedicines 10: 1803. https://doi.org/10.3390/biomedicines10081803
|
| [123] |
Murakami S, Mealey BL, Mariotti A, et al. (2018) Dental plaque-induced gingival conditions. J Clin Periodontol 45: S17-S27. https://doi.org/10.1002/JPER.17-0095
|
| [124] |
Van Dyke TE, Bgartold PM, Reynolds EC (2020) The nexus between periodontal inflammation and dysbiosis. Front Immunol 11: 511. https://doi.org/10.3389/fimmu.2020.00511
|
| [125] |
Abusleme L, Dupuy AK, Dutzan N, et al. (2013) The subgingival microbiome in health and periodontitis and its relationship with community biomass and inflammation. ISME J 7: 1016-1025. https://doi.org/10.1038/ismej.2012.174
|
| [126] |
Curtis MA, Diaz PI, Van Dyke TE (2020) The role of the microbiota in periodontal disease. Periodontol 83: 14-25. https://doi.org/10.1111/prd.12296
|
| [127] |
Schincaglia GP, Hong BY, Rosania A, et al. (2017) Clinical, immune, and microbiome traits of gingivitis and peri-implant mucositis. J Dent Res 96: 47-55. https://doi.org/10.1177/0022034516668847
|
| [128] |
Nowicki EM, Shroff R, Singleton JA, et al. (2018) Microbiota and metatranscriptome changes accompanying the onset of gingivitis. mBio 9: e00575-18. https://doi.org/10.1128/mBio.00575-18
|
| [129] |
Contaldo M, Itro A, Lajolo C, et al. (2020) Overview of osteoporosis, periodontitis and oral dysbiosis: The emerging role of oral microbiota. Appl Sci 10: 6000. https://doi.org/10.3390/app10176000
|
| [130] |
Scannapieco FA, Dongari-Bagtzoglou A (2021) Dysbiosis revisited: Understanding the role of the oral microbiome in the pathogenesis of gingivitis and periodontitis: A critical assessment. J Periodontol 92: 1071-1078. https://doi.org/10.1002/JPER.21-0120
|
| [131] |
Holmstrup P, Damgaard C, Olsen I, et al. (2017) Comorbidity of oeriodontal diseases: Two sides of the same coin?. J Oral Microbiol 9: 1332710. https://doi.org/10.1080/20002297.2017.1332710
|
| [132] |
Hajishengallis G, Chavakis T (2021) Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat Rev Immunol 21: 426-440. https://doi.org/10.1038/s41577-020-00488-6
|
| [133] |
Maitre Y, Mahalli R, Micheneau P, et al. (2021) Evidence and therapeutic perspectives in the relationship between the oral microbiome and Alzheimer's disease: A systematic review. Int J Environ Res Public Health 18: 11157. https://doi.org/10.3390/ijerph182111157
|
| [134] |
Poole S, Singhrao SK, Kesavalu L, et al. (2013) Determining the presence of periodontopathic virulence factors in short-term postmortem Alzheimer's disease brain tissue. J Alzheimers Dis 36: 665-677. https://doi.org/10.3233/JAD-121918
|
| [135] |
Vila T, Sultan AS, Montelongo-Jauregui D, et al. (2020) Oral candidiasis: A disease of opportunity. J Fungi 6: 15. https://doi.org/10.3390/jof6010015
|
| [136] |
Man A, Ciurea CN, Pasaroiu D, et al. (2017) New perspectives on the nutritional factors influencing growth rate of Candida albicans in diabetics. An in vitro study. Mem Inst Oswaldo Cruz 112: 587-592. https://doi.org/10.1590/0074-02760170098
|
| [137] | Boorghani M, Gholizadeh N, Taghavi Zenouz A, et al. (2010) Oral lichen planus: Clinical features, etiology, treatment and management; a review of literature. J Dent Res Dent Clin Dent Prospect 4: 3-9. https://doi.org/10.5681/joddd.2010.002 |
| [138] |
Bornstein MM, Hakimi B, Persson GRJ (2008) Microbiological findings in subjects with asymptomatic oral lichen planus: A cross-sectional comparative study. J Periodontol 79: 2347-2355. https://doi.org/10.1902/jop.2008.080303
|
| [139] |
Ertugrul AS, Arslan U, Dursun R, et al. (2013) Periodontopathogen profile of healthy and oral lichen planus patients with gingivitis or periodontitis. Int J Oral Sci 5: 92-97. https://doi.org/10.1038/ijos.2013.30
|
| [140] |
Choi YS, Kim Y, Yoon HJ, et al. (2016) The presence of bacteria within tissue provides insights into the pathogenesis of oral lichen planus. Sci Rep 6: 29186. https://doi.org/10.1038/srep29186
|
| [141] |
Yan L, Xu J, Lou F, et al. (2024) Alterations of oral microbiome and metabolic signatures and their interaction in oral lichen planus. J Oral Microbiol 16: 2422164. https://doi.org/10.1080/20002297.2024.2422164
|
| [142] |
Thomas C, Minty M, Vinel A, et al. (2021) Oral microbiota: A major player in the diagnosis of systemic diseases. Diagnostics 11: 1376. https://doi.org/10.3390/diagnostics11081376
|
| [143] |
Bourgeois D, Inquimbert C, Ottolenghi L, et al. (2019) Periodontal pathogens as risk factors of cardiovascular diseases, diabetes, rheumatoid arthritis, cancer, and chronic obstructive pulmonary disease-Is there cause for consideration?. Microorganisms 7: 424. https://doi.org/10.3390/microorganisms7100424
|
| [144] |
Chopra A, Franco-Duarte R, Rajagopal A, et al. (2024) Exploring the presence of oral bacteria in non-oral sites of patients with cardiovascular diseases using whole metagenomic data. Sci Rep 14: 1476. https://doi.org/10.1038/s41598-023-50891-x
|
| [145] |
Corrêa JD, Calderaro DC, Ferreira GA, et al. (2017) Subgingival microbiota dysbiosis in systemic lupus erythematosus: Association with periodontal status. Microbiome 5: 34. https://doi.org/10.1186/s40168-017-0252-z
|
| [146] | Yan X, Yang M, Liu J, et al. (2015) Discovery and validation of potential bacterial biomarkers for lung cancer. Am J Cancer Res 5: 3111-3122. |
| [147] |
Liu WJ, Xiao M, Yi J, et al. (2019) First case report of bacteremia caused by Solobacterium moorei in China, and literature review. BMC Infect Dis 19: 730. https://doi.org/10.1186/s12879-019-4359-7
|
| [148] |
Flemer B, Warren RD, Barrett MP, et al. (2018) The oral microbiota in colorectal cancer is distinctive and predictive. Gut 67: 1454-1463. https://doi.org/10.1136/gutjnl-2017-314814
|
| [149] |
Keogh RA, Doran KS (2023) Group B Streptococcus and diabetes: Finding the sweet spot. PLoS Pathog 19: e1011133. https://doi.org/10.1371/journal.ppat.1011133
|
| [150] |
Martini AM, Moricz BS, Ripperger AK, et al. (2020) Association of novel Streptococcus sanguinis virulence factors with pathogenesis in a native valve infective endocarditis model. Front Microbiol 11: 10. https://doi.org/10.3389/fmicb.2020.00010
|
| [151] |
Hammad MI, Conrads G, Abdelbary MMH (2023) Isolation, identification, and significance of salivary Veillonella spp., Prevotella spp., and Prevotella salivae in patients with inflammatory bowel disease. Front Cell Infect Microbiol 13: 1278582. https://doi.org/10.3389/fcimb.2023.1278582
|
| [152] | Saladi L, Zeana C, Singh M (2017) Native valve endocarditis due to Veillonella species: A case report and review of the literature. Case Rep Infect Dis 2017: 4896186. https://doi.org/10.1155/2017/4896186 |
| [153] |
Zhan Z, Liu W, Pan L, et al. (2022) Overabundance of Veillonella parvula promotes intestinal inflammation by activating macrophages via LPS-TLR4 pathway. Cell Death Discov 8: 251. https://doi.org/10.1038/s41420-022-01015-3
|
| [154] |
Ganesan SM, Joshi V, Fellows M, et al. (2017) A tale of two risks: Smoking, diabetes and the subgingival microbiome. ISME J 11: 2075-2089. https://doi.org/10.1038/ismej.2017.73
|
| [155] |
Eriksson K, Fei G, Lundmark A, et al. (2019) Periodontal health and oral microbiota in patients with rheumatoid arthritis. J Clin Med 8: 630. https://doi.org/10.3390/jcm8050630
|
| [156] |
Gnanasekaran J, Binder Gallimidi A, Saba E, et al. (2020) Intracellular Porphyromonas gingivalis promotes the tumorigenic behavior of pancreatic carcinoma cells. Cancers 12: 2331. https://doi.org/10.3390/cancers12082331
|
| [157] |
Huang X, Li Y, Zhang J, et al. (2024) Linking periodontitis with inflammatory bowel disease through the oral-gut axis: The potential role of Porphyromonas gingivalis. Biomedicines 12: 685. https://doi.org/10.3390/biomedicines12030685
|
| [158] |
Jia S, Li X, Du Q (2023) Host insulin resistance caused by Porphyromonas gingivalis - Review of recent progresses. Front Cell Infect Microbiol 13: 1209381. https://doi.org/10.3389/fcimb.2023.1209381
|
| [159] |
Li Y, Guo R, Oduro PK, et al. (2022) The relationship between Porphyromonas gingivalis and rheumatoid arthritis: A meta-analysis. Front Cell Infect Microbiol 12: 956417. https://doi.org/10.3389/fcimb.2022.956417
|
| [160] |
Malinowski B, Węsierska A, Zalewska K, et al. (2019) The role of Tannerella forsythia and Porphyromonas gingivalis in pathogenesis of esophageal cancer. Infect Agents Cancer 14: 3. https://doi.org/10.1186/s13027-019-0220-2
|
| [161] |
Reyes L, Phillips P, Wolfe B, et al. (2017) Porphyromonas gingivalis and adverse pregnancy outcome. J Oral Microbiol 10: 1374153. https://doi.org/10.1080/20002297.2017.1374153
|
| [162] |
Stanisic D, Jeremic N, Singh M, et al. (2023) Porphyromonas gingivalis induces cardiovascular dysfunction. Can J Physiol Pharmacol 101: 413-424. https://doi.org/10.1139/cjpp-2022-0392
|
| [163] |
Drago L (2019) Prevotella copri and microbiota in rheumatoid arthritis: Fully convincing evidence?. J Clin Med 8: 1837. https://doi.org/10.3390/jcm8111837
|
| [164] |
Guo J, Cui G, Huang W, et al. (2023) Alterations in the human oral microbiota in systemic lupus erythematosus. J Transl Med 21: 95. https://doi.org/10.1186/s12967-023-03892-3
|
| [165] |
Suzuki J, Imai Y, Aoki M, et al. (2014) Periodontitis in cardiovascular disease patients with or without Marfan syndrome-A possible role of Prevotella intermedia. PLoS One 9: e95521. https://doi.org/10.1371/journal.pone.0095521
|
| [166] |
Yuan H, Wu X, Wang X, et al. (2024) Microbial dysbiosis linked to metabolic dysfunction-associated fatty liver disease in Asians: Prevotella copri promotes lipopolysaccharide biosynthesis and network instability in the Prevotella enterotype. Int J Mol Sci 25: 2183. https://doi.org/10.3390/ijms25042183
|
| [167] |
Ardila CM, Perez-Valencia AY, Rendon-Osorio WL (2015) Tannerella forsythia is associated with increased levels of atherogenic low density lipoprotein and total cholesterol in chronic periodontitis. J Clin Expl Dent 7: e254-e260. https://doi.org/10.4317/jced.52128
|
| [168] |
Martínez-Rivera JI, Xibillé-Friedmann DX, González-Christen J, et al. (2017) Salivary ammonia levels and Tannerella forsythia are associated with rheumatoid arthritis: A cross sectional study. Clin Exp Dent Res 3: 107-114. https://doi.org/10.1002/cre2.68
|
| [169] | Schara R, Skaleric E, Seme K, et al. (2013) Prevalence of periodontal pathogens and metabolic control of type 1 diabetes patients. J Int Acad Periodontol 15: 29-34. |
| [170] | Dhaliwal D, Bhargava R, Movahed MR (2023) Fusobacterium nucleatum endocarditis: A case report and literature review. Am J Cardiovasc Dis 13: 29-31. |
| [171] |
Ghosh A, Jaaback K, Boulton A, et al. (2024) Fusobacterium nucleatum: An overview of evidence, demi-decadal trends, and its role in adverse pregnancy outcomes and various gynecological diseases, including cancers. Cells 13: 717. https://doi.org/10.3390/cells13080717
|
| [172] |
Luo L, Tang J, Du X, et al. (2024) Chronic obstructive pulmonary disease and the airway microbiome: A review for clinicians. Respir Med 225: 107586. https://doi.org/10.1016/j.rmed.2024.107586
|
| [173] |
Matsha T, Prince Y, Davids S, et al. (2020) Oral microbiome signatures in diabetes mellitus and periodontal disease. J Dent Res 99: 658-665. https://doi.org/10.1177/0022034520913818
|
| [174] |
Su W, Chen Y, Cao P, et al. (2020) Fusobacterium nucleatum promotes the development of ulcerative colitis by inducing the autophagic cell death of intestinal epithelial. Front Cell Infect Microbiol 10: 594806. https://doi.org/10.3389/fcimb.2020.594806
|
| [175] |
Zepeda-Rivera M, Minot SS, Bouzek H, et al. (2024) A distinct Fusobacterium nucleatum clade dominates the colorectal cancer niche. Nature 628: 424-432. https://doi.org/10.1038/s41586-024-07182-w
|
| [176] |
Castrillon CA, Hincapie JP, Yepes FL, et al. (2015) Occurrence of red complex microorganisms and Aggregatibacter actinomycetemcomitans in patients with diabetes. J Invest Clin Dent 6: 25-31. https://doi.org/10.1111/jicd.12051
|
| [177] |
Fan X, Alekseyenko AV, Wu J, et al. (2018) Human oral microbiome and prospective risk for pancreatic cancer: A population-based nested case-control study. Gut 67: 120-127. https://doi.org/10.1136/gutjnl-2016-312580
|
| [178] |
Santana PT, Rosas SLB, Ribeiro BE, et al. (2022) Dysbiosis in inflammatory bowel disease: Pathogenic role and potential therapeutic targets. Int J Mol Sci 23: 3464. https://doi.org/10.3390/ijms23073464
|
| [179] |
Talapko J, Juzbašić M, Meštrović T, et al. (2024) Aggregatibacter actinomycetemcomitans: From the oral cavity to the heart valves. Microorganisms 12: 1451. https://doi.org/10.3390/microorganisms12071451
|
| [180] |
Thomas C, Minty M, Canceill T, et al. (2021) Obesity drives an oral microbiota signature of female patients with periodontitis: A pilot study. Diagnostics 11: 745. https://doi.org/10.3390/diagnostics11050745
|
| [181] |
Zhang L, Lee H, Grimm MC, et al. (2014) Campylobacter concisus and inflammatory bowel disease. World J Gastroenterol 20: 1259-1267. https://doi.org/10.3748/wjg.v20.i5.1259
|
| [182] |
Short B, Carson S, Devlin AC, et al. (2021) Non-typeable Haemophilus influenzae chronic colonization in chronic obstructive pulmonary disease (COPD). Crit Rev Microbiol 47: 192-205. https://doi.org/10.1080/1040841X.2020.1863330
|
| [183] |
Murphy TF, Brauer AL, Pettigrew MM, et al. (2019) Persistence of Moraxella catarrhalis in chronic obstructive pulmonary disease and regulation of the Hag/MID adhesin. J Infect Dis 219: 1448-1455. https://doi.org/10.1093/infdis/jiy680
|
| [184] |
Borrego-Ruiz A, Borrego JJ (2024) An updated overview on the relationship between human gut microbiome dysbiosis and psychiatric and psychological disorders. Prog Neuropsychopharmacol Biol Psychiatry 128: 110861. https://doi.org/10.1016/j.pnpbp.2023.110861
|
| [185] |
Hashimoto K (2023) Emerging role of the host microbiome in neuropsychiatric disorders: Overview and future directions. Mol Psychiatry 28: 3625-3637. https://doi.org/10.1038/s41380-023-02287-6
|
| [186] |
Martin CR, Osadchiy V, Kalani A, et al. (2018) The brain-gut-microbiome axis. Cell Mol Gastroenterol Hepatol 6: 133-148. https://doi.org/10.1016/j.jcmgh.2018.04.003
|
| [187] |
Tiwari T, Kelly A, Randall CL, et al. (2022) Association between mental health and oral health status and care utilization. Front Oral Health 2: 732882. https://doi.org/10.3389/froh.2021.732882
|
| [188] |
Maitre Y, Micheneau P, Delpierre A, et al. (2020) Did the brain and oral microbiota talk to each other? A review of the literature. J Clin Med 9: 3876. https://doi.org/10.3390/jcm9123876
|
| [189] |
Skallevold HE, Rokaya N, Wongsirichat N, et al. (2023) Importance of oral health in mental health disorders: An updated review. J Oral Biol Craniofac Res 13: 544-552. https://doi.org/10.1016/j.jobcr.2023.06.003
|
| [190] |
Tao K, Yuan Y, Xie Q, et al. (2024) Relationship between human oral microbiome dysbiosis and neuropsychiatric diseases: An updated overview. Behav Brain Res 471: 115111. https://doi.org/10.1016/j.bbr.2024.115111
|
| [191] |
López-Valencia L, Moya M, Escudero B, et al. (2024) Bacterial lipopolysaccharide forms aggregates with apolipoproteins in male and female rat brains after ethanol binges. J Lipid Res 65: 100509. https://doi.org/10.1016/j.jlr.2024.100509
|
| [192] |
Yu XC, Yang JJ, Jin BH, et al. (2017) A strategy for bypassing the blood-brain barrier: Facial intradermal brain-targeted delivery via the trigeminal nerve. J Control Release 258: 22-33. https://doi.org/10.1016/j.jconrel.2017.05.001
|
| [193] |
Narengaowa, Kong W, Lan F, et al. (2021) The oral-gut-brain axis: The influence of microbes in Alzheimer's disease. Front Cell Neurosci 15: 633735. https://doi.org/10.3389/fncel.2021.633735
|
| [194] |
Martínez M, Martín-Hernández D, Virto L, et al. (2021) Periodontal diseases and depression: A pre-clinical in vivo study. J Clin Periodontol 48: 503-527. https://doi.org/10.1111/jcpe.13420
|
| [195] |
Ding Y, Ren J, Yu H, et al. (2018) Porphyromonas gingivalis, a periodontitis causing bacterium, induces memory impairment and age-dependent neuroinflammation in mice. Immun Ageing 15: 6. https://doi.org/10.1186/s12979-017-0110-7
|
| [196] |
Bowman GL, Dayon L, Kirkland R, et al. (2018) Blood-brain barrier breakdown, neuroinflammation, and cognitive decline in older adults. Alzheimers Dement 14: 1640-1650. https://doi.org/10.1016/j.jalz.2018.06.2857
|
| [197] |
Solár P, Zamani A, Kubíčková L, et al. (2020) Choroid plexus and the blood-cerebrospinal fluid barrier in disease. Fluids Barriers CNS 17: 35. https://doi.org/10.1186/s12987-020-00196-2
|
| [198] |
Bowland GB, Weyrich LS (2022) The oral-microbiome-brain axis and neuropsychiatric disorders: An anthropological perspective. Front Psychiatry 13: 810008. https://doi.org/10.3389/fpsyt.2022.810008
|
| [199] |
Bulgart HR, Neczypor EW, Wold LE, et al. (2020) Microbial involvement in Alzheimer disease development and progression. Mol Neurodegen 15: 42. https://doi.org/10.1186/s13024-020-00378-4
|
| [200] | Liu Y, Wu Z, Zhang X, et al. (2013) Leptomeningeal cells transduce peripheral macrophages inflammatory signal to microglia in reponse to Porphyromonas gingivalis LPS. Mediators Inflamm 2013: 407562. https://doi.org/10.1155/2013/407562 |
| [201] |
Sansores-España LD, Melgar-Rodríguez S, Olivares-Sagredo K, et al. (2021) Oral-gut-brain axis in experimental models of periodontitis: Associating gut dysbiosis with neurodegenerative diseases. Front Aging 2: 781582. https://doi.org/10.3389/fragi.2021.781582
|
| [202] |
Schmidt TS, Hayward MR, Coelho LP, et al. (2019) Extensive transmission of microbes along the gastrointestinal tract. eLife 8: e42693. https://doi.org/10.7554/eLife.42693
|
| [203] |
Atarashi K, Suda W, Luo C, et al. (2017) Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation. Science 358: 359-365. https://doi.org/10.1126/science.aan4526
|
| [204] |
Kitamoto S, Nagao-Kitamoto H, Hein R, et al. (2020) The bacterial connection between the oral cavity and the gut diseases. J Dent Res 99: 1021-1029. https://doi.org/10.1177/0022034520924633
|
| [205] |
Arimatsu K, Yamada H, Miyazawa H, et al. (2014) Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci Rep 4: 4828. https://doi.org/10.1038/srep04828
|
| [206] |
Feng YK, Wu QL, Peng YW, et al. (2020) Oral P. gingivalis impairs gut permeability and mediates immune responses associated with neurodegeneration in LRRK2 R1441G mice. J Neuroinflammation 17: 347. https://doi.org/10.1186/s12974-020-02027-5
|
| [207] |
Xue L, Zou X, Yang XQ, et al. (2020) Chronic periodontitis induces microbiota-gut-brain axis disorders and cognitive impairment in mice. Exp Neurol 326: 113176. https://doi.org/10.1016/j.expneurol.2020.113176
|
| [208] |
Liao C, Rolling T, Djukovic A, et al. (2024) Oral bacteria relative abundance in faeces increases due to gut microbiota depletion and is linked with patient outcomes. Nat Microbiol 9: 1555-1565. https://doi.org/10.1038/s41564-024-01680-3
|
| [209] |
Costa CFFA, Correia-de-Sá T, Araujo R, et al. (2024) The oral-gut microbiota relationship in healthy humans: Identifying shared bacteria between environments and age groups. Front Microbiol 15: 1475159. https://doi.org/10.3389/fmicb.2024.1475159
|
| [210] |
Kageyama S, Sakata S, Ma J, et al. (2023) High-resolution detection of translocation of oral bacteria to the gut. J Dent Res 102: 752-758. https://doi.org/10.1177/00220345231160747
|
| [211] |
Lu Y, Li Z, Peng X (2023) Regulatory effects of oral microbe on intestinal microbiota and the illness. Front Cell Infect Microbiol 13: 1093967. https://doi.org/10.3389/fcimb.2023.1093967
|
| [212] |
Borrego-Ruiz A, Borrego JJ (2024) Influence of human gut microbiome on the healthy and the neurodegenerative aging. Exp Gerontol 194: 112497. https://doi.org/10.1016/j.exger.2024.112497
|
| [213] |
Kamatham PT, Shukla R, Khatri DK, et al. (2024) Pathogenesis, diagnostics, and therapeutics for Alzheimer's disease: Breaking the memory barrier. Ageing Res Rev 101: 102481. https://doi.org/10.1016/j.arr.2024.102481
|
| [214] |
Chen GF, Xu TH, Yan Y, et al. (2017) Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol Sin 38: 1205-1235. https://doi.org/10.1038/aps.2017.28
|
| [215] |
Cerajewska TL, Davies M, West NX (2015) Periodontitis: A potential risk factor for Alzheimer's disease. Br Dent J 218: 29-34. https://doi.org/10.1038/sj.bdj.2014.1137
|
| [216] |
Tuganbaev T, Yoshida K, Honda K (2022) The effects of oral microbiota on health. Science 376: 934-936. https://doi.org/10.1126/science.abn1890
|
| [217] |
Wang RP, Ho YS, Leung WK, et al. (2019) Systemic inflammation linking chronic periodontitis to cognitive decline. Brain Behav Immun 81: 63-73. https://doi.org/10.1016/j.bbi.2019.07.002
|
| [218] |
Tzeng NS, Chung CH, Yeh CB, et al. (2016) Are chronic periodontitis and gingivitis associated with dementia? A nationwide, retrospective, matched-cohort study in Taiwan. Neuroepidemiology 47: 82-93. https://doi.org/10.1159/000449166
|
| [219] |
Chen CK, Wu YT, Chang YC (2017) Association between chronic periodontitis and the risk of Alzheimer's disease: A retrospective, population-based, matched-cohort study. Alzheimers Res Ther 9: 56. https://doi.org/10.1186/s13195-017-0282-6
|
| [220] |
Dominy SS, Lynch C, Ermini F, et al. (2019) Porphyromonas gingivalis in Alzheimer's disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci Adv 5: eaau3333. https://doi.org/10.1126/sciadv.aau3333
|
| [221] |
Kunkle BW, Grenier-Boley B, Sims R, et al. (2019) Genetic meta-analysis of diagnosed Alzheimer's disease identifies new risk loci and implicates Aβ, tau, immunity and lipid processing. Nat Genet 51: 414-430. https://doi.org/10.1038/s41588-019-0358-2
|
| [222] |
Kanagasingam S, von Ruhland C, Welbury R, et al. (2022) Porphyromonas gingivalis conditioned medium induces amyloidogenic processing of the amyloid-beta protein precursor upon in vitro infection of SH-SY5Y cells. J Alzheimers Dis Rep 6: 577-587. https://doi.org/10.3233/ADR-220029
|
| [223] |
Jin R, Ning X, Liu X, et al. (2023) Porphyromonas gingivalis-induced periodontitis could contribute to cognitive impairment in Sprague-Dawley rats via the P38 MAPK signaling pathway. Front Cell Neurosci 17: 1141339. https://doi.org/10.3389/fncel.2023.1141339
|
| [224] |
Morikawa T, Uehara O, Paudel D, et al. (2023) Systemic administration of lipopolysaccharide from Porphyromonas gingivalis decreases neprilysin expression in the mouse hippocampus. In Vivo 37: 163-172. https://doi.org/10.21873/invivo.13065
|
| [225] |
Carpentier M, Robitaille Y, DesGroseillers L, et al. (2002) Declining expression of neprilysin in Alzheimer disease vasculature: Possible involvement in cerebral amyloid angiopathy. J Neuropathol Exp Neurol 61: 849-856. https://doi.org/10.1093/jnen/61.10.849
|
| [226] |
Grimm MO, Mett J, Stahlmann CP, et al. (2013) Neprilysin and Aβ clearance: Impact of the APP intracellular domain in NEP regulation and implications in Alzheimer's disease. Front Aging Neurosci 5: 98. https://doi.org/10.3389/fnagi.2013.00098
|
| [227] |
Jungbauer G, Stahli A, Zhu X, et al. (2022) Periodontal microorganisms and Alzheimer disease - A causative relationship?. Periodontol 2000 89: 59-82. https://doi.org/10.1111/prd.12429
|
| [228] |
Díaz-Zúniga J, Munoz Y, Melgar-Rodríguez S, et al. (2019) Serotype b of Aggregatibacter actinomycetemcomitans triggers pro-inflammatory responses and amyloid beta secretion in hippocampal cells: A novel link between periodontitis and Alzheimeŕs disease?. J Oral Microbiol 11: 1586423. https://doi.org/10.1080/20002297.2019.1586423
|
| [229] |
Holt SC, Ebersole JL (2005) Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: The “red complex”, a prototype polybacterial pathogenic consortium in periodontitis. Periodontol 2000 38: 72-122. https://doi.org/10.1111/j.1600-0757.2005.00113.x
|
| [230] |
Riviere GR, Riviere KH, Smith KS (2002) Molecular and immunological evidence of oral Treponema in the human brain and their association with Alzheimer's disease. Oral Microbiol Immunol 17: 113-118. https://doi.org/10.1046/j.0902-0055.2001.00100.x
|
| [231] |
Su X, Tang Z, Lu Z, et al. (2021) Oral Treponema denticola infection induces Aβ1-40 and Aβ1-42 accumulation in the hippocampus of C57BL/6 mice. J Mol Neurosci 71: 1506-1514. https://doi.org/10.1007/s12031-021-01827-5
|
| [232] |
Wu L, Su X, Tang Z, et al. (2022) Treponema denticola induces neuronal apoptosis by promoting amyloid-beta accumulation in mice. Pathogens 11: 1150. https://doi.org/10.3390/pathogens11101150
|
| [233] |
Yan C, Diao Q, Zhao Y, et al. (2022) Fusobacterium nucleatum infection-induced neurodegeneration and abnormal gut microbiota composition in Alzheimer's disease-like rats. Front Neurosci 16: 884543. https://doi.org/10.3389/fnins.2022.884543
|
| [234] |
Wan J, Fan H (2023) Oral microbiome and Alzheimer's disease. Microorganisms 11: 2550. https://doi.org/10.3390/microorganisms11102550
|
| [235] |
Watanabe I, Kuriyama N, Miyatani F, et al. (2016) Oral Cnm-positive Streptococcus mutans expressing collagen binding activity is a risk factor for cerebral microbleeds and cognitive impairment. Sci Rep 6: 38561. https://doi.org/10.1038/srep38561
|
| [236] |
Wang DN, Hou XW, Yang BW, et al. (2015) Quantity of cerebral microbleeds, antiplatelet therapy, and intracerebral hemorrhage outcomes: A systematic review and meta-analysis. J Stroke Cerebrovasc Dis 24: 2728-2737. https://doi.org/10.1016/j.jstrokecerebrovasdis.2015.08.003
|
| [237] |
Liu XX, Jiao B, Liao XX, et al. (2019) Analysis of salivary microbiome in patients with Alzheimer's disease. J Alzheimers Dis 72: 633-640. https://doi.org/10.3233/JAD-190587
|
| [238] |
Wu YF, Lee WF, Salamanca E, et al. (2021) Oral microbiota changes in elderly patients, an indicator of Alzheimer's disease. Int J Environ Res Public Health 18: 4211. https://doi.org/10.3390/ijerph18084211
|
| [239] |
Cirstea MS, Kliger D, MacLellan AD, et al. (2022) The oral and fecal microbiota in a Canadian cohort of Alzheimer's disease. J Alzheimers Dis 87: 247-258. https://doi.org/10.3233/JAD-215520
|
| [240] |
Fu KL, Chiu MJ, Wara-Aswapati N, et al. (2023) Oral microbiome and serological analyses on association of Alzheimer's disease and periodontitis. Oral Dis 29: 3677-3687. https://doi.org/10.1111/odi.14348
|
| [241] |
Issilbayeva A, Kaiyrlykyzy A, Vinogradova E, et al. (2024) Oral microbiome stamp in Alzheimer's disease. Pathogens 13: 195. https://doi.org/10.3390/pathogens13030195
|
| [242] |
Holmer J, Aho V, Eriksdotter M, et al. (2021) Subgingival microbiota in a population with and without cognitive dysfunction. J Oral Microbiol 13: 1854552. https://doi.org/10.1080/20002297.2020.1854552
|
| [243] |
Jankovic J, Tan EK (2020) Parkinson's disease: Etiopathogenesis and treatment. J Neurol Neurosurg Psychiatry 91: 795-808. https://doi.org/10.1136/jnnp-2019-322338
|
| [244] |
Fleury V, Zekeridou A, Lazarevic V, et al. (2021) Oral dysbiosis and inflammation in Parkinson's disease. J Parkinsons Dis 11: 619-631. https://doi.org/10.3233/JPD-202459
|
| [245] |
Rozas NS, Tribble GD, Jeter CB (2021) Oral factors that impact the oral microbiota in Parkinson's disease. Microorganisms 9: 1616. https://doi.org/10.3390/microorganisms9081616
|
| [246] |
Bai XB, Xu S, Zhou LJ, et al. (2023) Oral pathogens exacerbate Parkinson's disease by promoting Th1 cell infiltration in mice. Microbiome 11: 254. https://doi.org/10.1186/s40168-023-01685-w
|
| [247] |
Jaber MA (2011) Dental caries experience, oral health status and treatment needs of dental patients with autism. J Appl Oral Sci 19: 212-217. https://doi.org/10.1590/s1678-77572011000300006
|
| [248] | Ferrazzano GF, Salerno C, Bravaccio C, et al. (2020) Autism spectrum disorders and oral health status: Review of the literature. Eur J Paediatr Dent 21: 9-12. https://doi.org/10.23804/ejpd.2020.21.01.02 |
| [249] |
Mussap M, Beretta P, Esposito E, et al. (2023) Once upon a time oral microbiota: A cinderella or a protagonist in Autism Spectrum Disorder?. Metabolites 13: 1183. https://doi.org/10.3390/metabo13121183
|
| [250] |
Olsen I, Hicks SD (2019) Oral microbiota and autism spectrum disorder (ASD). J Oral Microbiol 12: 1702806. https://doi.org/10.1080/20002297.2019.1702806
|
| [251] |
Kealy J, Greene C, Campbell M (2020) Blood-brain barrier regulation in psychiatric disorders. Neurosci Lett 726: 133664. https://doi.org/10.1016/j.neulet.2018.06.033
|
| [252] |
Srikantha P, Mohajeri MH (2019) The possible role of the microbiota-gut-brain-axis in autism spectrum disorder. Int J Mol Sci 20: 2115. https://doi.org/10.3390/ijms20092115
|
| [253] |
Tedjosasongko U, Oktaviani PD, Nadia S, et al. (2023) Can oral microbiome dysbiosis affect the behavior of children with Autism Spectrum Disorders (ASD)?: Narrative review. World J Adv Res Rev 17: 51-56. https://doi.org/10.30574/wjarr.2023.17.1.1461
|
| [254] |
Johnson D, Letchumanan V, Thurairajasingam S, et al. (2020) A revolutionizing approach to autism spectrum disorder using the microbiome. Nutrients 12: 1983. https://doi.org/10.3390/nu12071983
|
| [255] |
Hicks SD, Uhlig R, Afshari P, et al. (2018) Oral microbiome activity in children with autism spectrum disorder. Autism Res 11: 1286-1299. https://doi.org/10.1002/aur.1972
|
| [256] |
Sivamaruthi BS, Suganthy N, Kesika P, et al. (2020) The role of microbiome, dietary supplements, and probiotics in autism spectrum disorder. Int J Environ Res Public Health 17: 2647. https://doi.org/10.3390/ijerph17082647
|
| [257] |
Qiao Y, Wu M, Feng Y, et al. (2018) Alterations of oral microbiota distinguish children with autism spectrum disorders from healthy controls. Sci Rep 8: 1597. https://doi.org/10.1038/s41598-018-19982-y
|
| [258] |
Kong X, Liu J, Cetinbas M, et al. (2019) New and preliminary evidence on altered oral and gut microbiota in individuals with Autism Spectrum Disorder (ASD): Implications for ASD diagnosis and subtyping based on microbial biomarkers. Nutrients 11: 2128. https://doi.org/10.3390/nu11092128
|
| [259] |
Abdulhaq A, Halboub E, Homeida HE, et al. (2021) Tongue microbiome in children with autism spectrum disorder. J Oral Microbiol 13: 1936434. https://doi.org/10.1080/20002297.2021.1936434
|
| [260] |
Manghi P, Filosi M, Zolfo M, et al. (2024) Large-scale metagenomic analysis of oral microbiomes reveals markers for autism spectrum disorders. Nat Commun 15: 9743. https://doi.org/10.1038/s41467-024-53934-7
|
| [261] |
Garbarino VR, Gilman TL, Daws LC (2019) Extreme enhancement or depletion of serotonin transporter function and serotonin availability in autism spectrum disorder. Pharmacol Res 140: 85-99. https://doi.org/10.1016/j.phrs.2018.07.010
|
| [262] |
Evenepoel M, Daniels N, Moerkerke M, et al. (2024) Oral microbiota in autistic children: Diagnosis-related differences and associations with clinical characteristics. Brain Behav Immun Health 38: 100801. https://doi.org/10.1016/j.bbih.2024.100801
|
| [263] |
Szuhany KL, Simon NM (2022) Anxiety disorders: A review. JAMA 328: 2431-2445. https://doi.org/10.1001/jama.2022.22744
|
| [264] |
Johnson PL, Federici LM, Shekhar A (2014) Etiology, triggers and neurochemical circuits associated with unexpected, expected, and laboratory-induced panic attacks. Neurosci Biobehav Rev 46: 429-454. https://doi.org/10.1016/j.neubiorev.2014.07.027
|
| [265] |
Krupa-Kotara K, Gwioździk W, Nandzik S, et al. (2023) The role of microbiota pattern in anxiety and stress disorders-A review of the state of knowledge. Psych 5: 602-618. https://doi.org/10.3390/psych5030038
|
| [266] |
Nasir M, Trujillo D, Levine J, et al. (2020) Glutamate systems in DSM-5 anxiety disorders: Their role and a review of glutamate and GABA psychopharmacology. Front Psychiatry 11: 548505. https://doi.org/10.3389/fpsyt.2020.548505
|
| [267] |
Hsu CC, Hsu YC, Chen HJ, et al. (2015) Association of periodontitis and subsequent depression: A nationwide population-based study. Medicine 94: e2347. https://doi.org/10.1097/MD.0000000000002347
|
| [268] |
Malan-Müller S, Vidal R, O'Shea E, et al. (2024) Probing the oral-brain connection: Oral microbiome patterns in a large community cohort with anxiety, depression, and trauma symptoms, and periodontal outcomes. Transl Psychiatry 14: 419. https://doi.org/10.1038/s41398-024-03122-4
|
| [269] |
Martínez M, Postolache TT, García-Bueno B, et al. (2022) The role of the oral microbiota related to periodontal diseases in anxiety, mood and trauma- and stress-related disorders. Front Psychiatry 12: 814177. https://doi.org/10.3389/fpsyt.2021.814177
|
| [270] |
Xie Z, Jiang W, Deng M, et al. (2021) Alterations of oral microbiota in patients with panic disorder. Bioengineered 12: 9103-9112. https://doi.org/10.1080/21655979.2021.1994738
|
| [271] |
Faravelli C, Lo Sauro C, Lelli L, et al. (2012) The role of life events and HPA axis in anxiety disorders: A review. Curr Pharm Des 18: 5663-5674. https://doi.org/10.2174/138161212803530907
|
| [272] |
Juruena MF, Eror F, Cleare AJ, et al. (2020) The role of early life stress in HPA axis and anxiety. Adv Exp Med Biol 1191: 141-153. https://doi.org/10.1007/978-981-32-9705-0_9
|
| [273] |
Simpson CA, Adler C, du Plessis MR, et al. (2020) Oral microbiome composition, but not diversity, is associated with adolescent anxiety and depression symptoms. Physiol Behav 226: 113126. https://doi.org/10.1016/j.physbeh.2020.113126
|
| [274] |
Malhi GS, Mann JJ (2018) Depression. Lancet 392: 2299-2312. https://doi.org/10.1016/S0140-6736(18)31948-2
|
| [275] | Bernard JER (2018) Depression: A review of its definition. MOJ Addict Med Ther 5: 6-7. https://doi.org/10.15406/mojamt.2018.05.00082 |
| [276] |
Serafini RA, Pryce KD, Zachariou V (2020) The mesolimbic dopamine system in chronic pain and associated affective comorbidities. Biol Psychiatry 87: 64-73. https://doi.org/10.1016/j.biopsych.2019.10.018
|
| [277] |
Moncrieff J, Cooper RE, Stockmann T, et al. (2023) The serotonin theory of depression: A systematic umbrella review of the evidence. Mol Psychiatry 28: 3243-3256. https://doi.org/10.1038/s41380-022-01661-0
|
| [278] |
Daut RA, Fonken LK (2019) Circadian regulation of depression: A role for serotonin. Front Neuroendocrinol 54: 100746. https://doi.org/10.1016/j.yfrne.2019.04.003
|
| [279] |
Peng GJ, Tian JS, Gao XX, et al. (2015) Research on the pathological mechanism and drug treatment mechanism of depression. Curr Neuropharmacol 13: 514-523. https://doi.org/10.2174/1570159x1304150831120428
|
| [280] |
Johannsen A, Rylander G, Soder B, et al. (2006) Dental plaque, gingival inflammation, and elevated levels of interleukin-6 and cortisol in gingival crevicular fluid from women with stress-related depression and exhaustion. J Periodontol 77: 1403-1409. https://doi.org/10.1902/jop.2006.050411
|
| [281] |
Duran-Pinedo AE, Solbiati J, Frias-Lopez J (2018) The effect of the stress hormone cortisol on the metatranscriptome of the oral microbiome. Npj Biofilms Microbiomes 4: 25. https://doi.org/10.1038/s41522-018-0068-z
|
| [282] |
Wingfield B, Lapsley C, McDowell A, et al. (2021) Variations in the oral microbiome are associated with depression in young adults. Sci Rep 11: 15009. https://doi.org/10.1038/s41598-021-94498-6
|
| [283] |
Al Bataineh MT, Künstner A, Dash NR, et al. (2022) Altered composition of the oral microbiota in depression among cigarette smokers: A pilot study. Front Psychiatry 13: 902433. https://doi.org/10.3389/fpsyt.2022.902433
|
| [284] |
Li C, Chen Y, Wen Y, et al. (2022) A genetic association study reveals the relationship between the oral microbiome and anxiety and depression symptoms. Front Psychiatry 13: 960756. https://doi.org/10.3389/fpsyt.2022.960756
|
| [285] |
Alotiby A (2024) Immunology of stress: A review article. J Clin Med 13: 6394. https://doi.org/10.3390/jcm13216394
|
| [286] |
Paudel D, Uehara O, Giri S, et al. (2022) Effect of psychological stress on the oral-gut microbiota and the potential oral-gut-brain axis. Jpn Dent Sci Rev 58: 365-375. https://doi.org/10.1016/j.jdsr.2022.11.003
|
| [287] | Yaribeygi H, Panahi Y, Sahraei H, et al. (2017) The impact of stress on body function: A review. EXCLI J 16: 1057-1072. https://doi.org/10.17179/excli2017-480 |
| [288] |
Sonnenburg JL, Sonnenburg ED (2019) Vulnerability of the industrialized microbiota. Science 366: eaaw9255. https://doi.org/10.1126/science.aaw9255
|
| [289] |
Godoy LD, Rossignoli MT, Delfino-Pereira P, et al. (2018) A comprehensive overview on stress neurobiology: Basic concepts and clinical implications. Front Behav Neurosci 12: 127. https://doi.org/10.3389/fnbeh.2018.00127
|
| [290] |
Cain DW, Cidlowski JA (2017) Immune regulation by glucocorticoids. Nat Rev Immunol 17: 233-247. https://doi.org/10.1038/nri.2017.1
|
| [291] |
Weinstein LI, Revuelta A, Pando RH (2015) Catecholamines and acetylcholine are key regulators of the interaction between microbes and the immune system. Ann N Y Acad Sci 1351: 39-51. https://doi.org/10.1111/nyas.12792
|
| [292] |
Herman JP, McKlveen JM, Ghosal S, et al. (2016) Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr Physiol 6: 603-621. https://doi.org/10.1002/cphy.c150015
|
| [293] |
Agorastos A, Chrousos GP (2022) The neuroendocrinology of stress: The stress-related continuum of chronic disease development. Mol Psychiatry 27: 502-513. https://doi.org/10.1038/s41380-021-01224-9
|
| [294] |
Elenkov IJ (2002) Systemic stress-induced Th2 shift and its clinical implications. Int Rev Neurobiol 52: 163-186. https://doi.org/10.1016/s0074-7742(02)52009-2
|
| [295] |
Par M, Tarle Z (2019) Psychoneuroimmunology of oral diseases-A review. Stomatol Edu J 6: 55-65. https://doi.org/10.25241/stomaeduj.2019.6(1).art.7
|
| [296] |
Ganesan A, Kumar G, Gauthaman J, et al. (2024) Exploring the relationship between psychoneuroimmunology and oral diseases: A comprehensive review and analysis. J Lifestyle Med 14: 13-19. https://doi.org/10.15280/jlm.2024.14.1.13
|
| [297] |
Engeland CG, Bosch JA, Rohleder N (2019) Salivary biomarkers in psychoneuroimmunology. Curr Opin Behav Sci 28: 58-65. https://doi.org/10.1016/j.cobeha.2019.01.007
|
| [298] |
Zhang J, Lin S, Luo L, et al. (2023) Psychological stress: Neuroimmune roles in periodontal disease. Odontology 111: 554-564. https://doi.org/10.1007/s10266-022-00768-8
|
| [299] |
Roberts A, Matthews JB, Socransky SS, et al. (2002) Stress and the periodontal diseases: Effects of catecholamines on the growth of periodontal bacteria in vitro. Oral Microbiol Immunol 17: 296-303. https://doi.org/10.1034/j.1399-302X.2002.170506.x
|
| [300] |
Calil CM, Oliveira GM, Cogo K, et al. (2014) Effects of stress hormones on the production of volatile sulfur compounds by periodontopathogenic bacteria. Braz Oral Res 28: S1806-83242014000100228. https://doi.org/10.1590/1807-3107bor-2014.vol28.0008
|
| [301] |
de Lima PO, Nani BD, Almeida B, et al. (2018) Stress-related salivary proteins affect the production of volatile sulfur compounds by oral bacteria. Oral Dis 24: 1358-1366. https://doi.org/10.1111/odi.12890
|
| [302] |
Nani BD, Lima PO, Marcondes FK, et al. (2017) Changes in salivary microbiota increase volatile sulfur compounds production in healthy male subjects with academic-related chronic stress. PLoS One 12: e0173686. https://doi.org/10.1371/journal.pone.0173686
|
| [303] |
Kohn JN, Kosciolek T, Marotz C, et al. (2020) Differing salivary microbiome diversity, community and diurnal rhythmicity in association with affective state and peripheral inflammation in adults. Brain Behav Immun 87: 591-602. https://doi.org/10.1016/j.bbi.2020.02.004
|
| [304] |
Levert-Levitt E, Shapira G, Sragovich S, et al. (2022) Oral microbiota signatures in post-traumatic stress disorder (PTSD) veterans. Mol Psychiatry 27: 4590-4598. https://doi.org/10.1038/s41380-022-01704-6
|
| [305] |
Stoy S, McMillan A, Ericsson AC, et al. (2023) The effect of physical and psychological stress on the oral microbiome. Front Psychol 14: 1166168. https://doi.org/10.3389/fpsyg.2023.1166168
|
| [306] |
Alex AM, Levendosky AA, Bogat GA, et al. (2024) Stress and mental health symptoms in early pregnancy are associated with the oral microbiome. BMJ Ment Health 27: e301100. https://doi.org/10.1136/bmjment-2024-301100
|
| [307] |
Charalambous EG, Mériaux SB, Guebels P, et al. (2024) The oral microbiome is associated with HPA axis response to a psychosocial stressor. Sci Rep 14: 15841. https://doi.org/10.1038/s41598-024-66796-2
|
| [308] |
Akcali A, Huck O, Tenenbaum H, et al. (2013) Periodontal diseases and stress: A brief review. J Oral Rehabil 40: 60-68. https://doi.org/10.1111/j.1365-2842.2012.02341.x
|
| [309] |
Kim HM, Rothenberger CM, Davey ME (2022) Cortisol promotes surface translocation of Porphyromonas gingivalis. Pathogens 11: 982. https://doi.org/10.3390/pathogens11090982
|
| [310] |
Jentsch HF, März D, Krüger M (2013) The effects of stress hormones on growth of selected periodontitis related bacteria. Anaerobe 24: 49-54. https://doi.org/10.1016/j.anaerobe.2013.09.001
|
| [311] |
Zhang Y, Chen R, Zhang D, et al. (2023) Metabolite interactions between host and microbiota during health and disease: Which feeds the other?. Biomed Pharmacother 160: 114295. https://doi.org/10.1016/j.biopha.2023.114295
|
| [312] |
McIntyre RS, Calabrese JR (2019) Bipolar depression: The clinical characteristics and unmet needs of a complex disorder. Curr Med Res Opin 35: 1993-2005. https://doi.org/10.1080/03007995.2019.1636017
|
| [313] |
Baek JH, Jung SJ, Peters A, et al. (2023) Association between periodontal diseases and bipolar disorder: Implications for therapeutic interventions: A narrative review. Precis Future Med 7: 117-122. https://doi.org/10.23838/pfm.2023.00086
|
| [314] |
Cunha FA, Cota LOM, Cortelli SC, et al. (2018) Periodontal condition and levels of bacteria associated with periodontitis in individuals with bipolar affective disorders: A case-control study. J Periodontal Res 54: 63-72. https://doi.org/10.1111/jre.12605
|
| [315] |
Stępnicki P, Kondej M, Kaczor AA (2018) Current concepts and treatments of schizophrenia. Molecules 23: 2087. https://doi.org/10.3390/molecules23082087
|
| [316] |
Martin S, Foulon A, El Hage W, et al. (2022) Is there a link between oropharyngeal microbiome and schizophrenia? A narrative review. Int J Mol Sci 23: 846. https://doi.org/10.3390/ijms23020846
|
| [317] |
Murray N, Al Khalaf S, Kaulmann D, et al. (2021) Compositional and functional alterations in the oral and gut microbiota in patients with psychosis or schizophrenia: A systematic review. HRB Open Res 4: 108. https://doi.org/10.12688/hrbopenres.13416.1
|
| [318] |
Cui G, Qing Y, Li M, et al. (2021) Salivary metabolomics reveals that metabolic alterations precede the onset of schizophrenia. J Proteome Res 20: 5010-5023. https://doi.org/10.1021/acs.jproteome.1c00504
|
| [319] |
Castro-Nallar E, Bendall ML, Pérez-Losada M, et al. (2015) Composition, taxonomy and functional diversity of the oropharynx microbiome in individuals with schizophrenia and controls. PeerJ 3: e1140. https://doi.org/10.7717/peerj.1140
|
| [320] |
Dickerson F, Severance E, Yolken R (2017) The microbiome, immunity, and schizophrenia and bipolar disorder. Brain Behav Immun 62: 46-52. https://doi.org/10.1016/j.bbi.2016.12.010
|
| [321] |
Yolken R, Prandovszky E, Severance EG, et al. (2021) The oropharyngeal microbiome is altered in individuals with schizophrenia and mania. Schizophr Res 234: 51-57. https://doi.org/10.1016/j.schres.2020.03.010
|
| [322] |
Qing Y, Xu L, Cui G, et al. (2021) Salivary microbiome profiling reveals a dysbiotic schizophrenia-associated microbiota. NPJ Schizophr 7: 51. https://doi.org/10.1038/s41537-021-00180-1
|
| [323] |
Ling Z, Cheng Y, Liu X, et al. (2023) Altered oral microbiota and immune dysfunction in Chinese elderly patients with schizophrenia: A cross-sectional study. Transl Psychiatry 13: 383. https://doi.org/10.1038/s41398-023-02682-1
|
| [324] |
Liu X, Ling Z, Cheng Y, et al. (2024) Oral fungal dysbiosis and systemic immune dysfunction in Chinese patients with schizophrenia. Transl Psychiatry 14: 475. https://doi.org/10.1038/s41398-024-03183-5
|
| [325] |
Botero JE, Rodríguez-Medina C, Amaya-Sanchez S, et al. (2024) A comprehensive review of the relationship between oral health and Down syndrome. Curr Oral Health Rep 11: 15-22. https://doi.org/10.1007/s40496-024-00363-6
|
| [326] | Reuland-Bosma W, Van der Reijden WA, VanWinkelhoff AJ (2001) Absence of a specific subgingival microflora in adults with Down's syndrome. J Clin Periodontol 28: 1004-1009. https://doi.org/10.1034/j.1600-051x.2001.281103.x |
| [327] |
Amano A, Kishima T, Akiyama S, et al. (2001) Relationship of periodontopathic bacteria with early-onset periodontitis in Down's syndrome. J Periodontol 72: 368-373. https://doi.org/10.1902/jop.2001.72.3.368
|
| [328] | Linossier AG, Valenzuela CY, Toledo H (2008) Differences of the oral colonization by Streptococcus of the mutans group in children and adolescents with Down syndrome, mental retardation and normal controls. Med Oral Patol Oral Cir Bucal 13: 536-539. |
| [329] |
Khocht A, Yaskell T, Janal M (2012) Subgingival microbiota in adult down syndrome periodontitis. J Periodontal Res 47: 500-507. https://doi.org/10.1111/j.1600-0765.2011.01459.x
|
| [330] |
Willis JR, Iraola-Guzmán S, Saus E, et al. (2020) Oral microbiome in down syndrome and its implications on oral health. J Oral Microbiol 13: 1865690. https://doi.org/10.1080/20002297.2020.1865690
|
| [331] |
Cuenca M, Marín MJ, Nóvoa L, et al. (2021) Periodontal condition and subgingival microbiota characterization in subjects with Down syndrome. Appl Sci 11: 778. https://doi.org/10.3390/app11020778
|
| [332] |
Mitsuhata C, Kado N, Hamada M, et al. (2022) Characterization of the unique oral microbiome of children with Down syndrome. Sci Rep 12: 14150. https://doi.org/10.1038/s41598-022-18409-z
|
| [333] |
Contaldo M, Lucchese A, Romano A, et al. (2021) Oral microbiota features in subjects with Down syndrome and periodontal diseases: A systematic review. Int J Mol Sci 22: 9251. https://doi.org/10.3390/ijms22179251
|
| [334] |
Nóvoa Garrido L, Sánchez MD, Blanco Carrión J, et al. (2020) The subgingival microbiome in patients with Down syndrome and periodontitis. J Clin Med 9: 2482. https://doi.org/10.3390/jcm9082482
|
| [335] | Diéguez-Pérez M, de Nova-García MJ, Mourelle-Martínez MR, et al. (2016) Oral health in children with physical (Cerebral Palsy) and intellectual (Down Syndrome) disabilities: Systematic review I. J Clin Exp Dent 8: e337-e343. https://doi.org/10.4317/jced.52922 |
| [336] |
Sinha N, Singh B, Chhabra KG, et al. (2015) Comparison of oral health status between children with cerebral palsy and normal children in India: A case-control study. J Indian Soc Periodontol 19: 78-82. https://doi.org/10.4103/0972-124X.145800
|
| [337] |
Huang C, Chu C, Peng Y, et al. (2022) Correlations between gastrointestinal and oral microbiota in children with cerebral palsy and epilepsy. Front Pediatr 10: 988601. https://doi.org/10.3389/fped.2022.988601
|
| [338] |
Liu M, Shi Y, Wu K, et al. (2022) From mouth to brain: Distinct supragingival plaque microbiota composition in cerebral palsy children with caries. Front Cell Infect Microbiol 12: 814473. https://doi.org/10.3389/fcimb.2022.814473
|
| [339] |
Lian X, Liu Z, Wu T, et al. (2023) Oral microbiome alterations in epilepsy and after seizure control. Front Microbiol 14: 1277022. https://doi.org/10.3389/fmicb.2023.1277022
|
| [340] |
Zhao C, Chen F, Li Q, et al. (2024) Causal relationship between oral microbiota and epilepsy risk: Evidence from Mendelian randomization analysis in East Asians. Epilepsia Open 9: 2419-2428. https://doi.org/10.1002/epi4.13074
|
| [341] | World Health OrganizationOral health (2024). Available online: https://Www.Who.Int/Health-Topics/Oral-Health/#tab=tab_1 (accessed on 4 December 2024) |
| [342] |
Varoni EM, Rimondini L (2022) Oral microbiome, oral health and systemic health: A multidirectional link. Biomedicines 10: 186. https://doi.org/10.3390/biomedicines10010186
|
| [343] |
Militi A, Sicari F, Portelli M, et al. (2021) Psychological and social effects of oral health and dental aesthetic in adolescence and early adulthood: An observational study. Int J Environ Res Public Health 18: 9022. https://doi.org/10.3390/ijerph18179022
|
| [344] |
Ferreira MC, Dias-Pereira AC, Branco-de-Almeida LS, et al. (2017) Impact of periodontal disease on quality of life: A systematic review. J Periodontal Res 52: 651-665. https://doi.org/10.1111/jre.12436
|
| [345] |
Kastenbom L, Falsen A, Larsson P, et al. (2019) Costs and health-related quality of life in relation to caries. BMC Oral Health 19: 187. https://doi.org/10.1186/s12903-019-0874-6
|
| [346] |
Yuwanati M, Gondivkar S, Sarode SC, et al. (2021) Oral health-related quality of life in oral cancer patients: Systematic review and meta-analysis. Future Oncol 17: 979-990. https://doi.org/10.2217/fon-2020-0881
|
| [347] |
Wensel CR, Pluznick JL, Salzberg SL, et al. (2022) Next-generation sequencing: Insights to advance clinical investigations of the microbiome. J Clinical Invest 132: e154944. https://doi.org/10.1172/JCI154944
|
| [348] |
Nomura Y, Inai Y, Okada A, et al. (2023) Oral microbiome and oral health: Current stage and future prospective. Appl Sci 13: 11372. https://doi.org/10.3390/app132011372
|
| [349] | Chi L, Cheng X, Lin L, et al. (2021) Porphyromonas gingivalis-induced cognitive impairment is associated with gut dysbiosis, neuroinflammation, and glymphatic dysfunction. Front Cell Infect Microbiol 11: 977. https://doi.org/10.3389/fcimb.2021.755925 |
| [350] |
Simas AM, Kramer CD, Weinberg EO, et al. (2021) Oral infection with a periodontal pathogen alters oral and gut microbiomes. Anaerobe 71: 102399. https://doi.org/10.1016/j.anaerobe.2021.102399
|
| [351] |
Kato T, Yamazaki K, Nakajima M, et al. (2018) Oral administration of Porphyromonas gingivalis alters the gut microbiome and serum metabolome. mSphere 3: e00460-18. https://doi.org/10.1128/mSphere.00460-18
|
| [352] |
Leblhuber F, Ehrlich D, Steiner K, et al. (2021) The immunopathogenesis of Alzheimer's disease is related to the composition of gut microbiota. Nutrients 13: 361. https://doi.org/10.3390/nu13020361
|
| [353] |
Preshaw PM (2018) Host modulation therapy with anti-inflammatory agents. Periodontology 2000 76: 131-149. https://doi.org/10.1111/prd.12148
|
| [354] |
Yurko-Mauro K, McCarthy D, Rom D, et al. (2010) Beneficial effects of docosahexaenoic acid on cognition in age-related cognitive decline. Alzheimers Dement 6: 456-464. https://doi.org/10.1016/j.jalz.2010.01.013
|
| [355] |
Borrego-Ruiz A, González-Domenech CM, Borrego JJ (2024) The role of fermented vegetables as a sustainable and health-promoting nutritional resource. Appl Sci 14: 10853. https://doi.org/10.3390/app142310853
|
| [356] |
Ribeiro-Vidal H, Sánchez MC, Alonso-Español A, et al. (2020) Antimicrobial activity of EPA and DHA against oral pathogenic bacteria using an in vitro multi-species subgingival biofilm model. Nutrients 12: 2812. https://doi.org/10.3390/nu12092812
|
| [357] |
Khalifa K, Bergland AK, Soennesyn H, et al. (2020) Effects of purified anthocyanins in people at risk for dementia: Study protocol for a phase II randomized controlled trial. Front Neurol 11: 916. https://doi.org/10.3389/fneur.2020.00916
|
| [358] |
Ullah R, Khan M, Shah SA, et al. (2019) Natural antioxidant anthocyanins - A hidden therapeutic candidate in metabolic disorders with major focus in neurodegeneration. Nutrients 11: 1195. https://doi.org/10.3390/nu11061195
|
| [359] |
Guo Y, Li Z, Chen F, et al. (2023) Polyphenols in oral health: Homeostasis maintenance, disease prevention, and therapeutic applications. Nutrients 15: 4384. https://doi.org/10.3390/nu15204384
|
| [360] |
Kovac J, Slobodnikova L, Trajcikova E, et al. (2022) Therapeutic potential of flavonoids and tannins in management of oral infectious diseases - A review. Molecules 28: 158. https://doi.org/10.3390/molecules28010158
|
| [361] |
Gorr SU, Abdolhosseini M (2011) Antimicrobial peptides and periodontal disease. J Clin Periodontol 38: 126-141. https://doi.org/10.1111/j.1600-051X.2010.01664.x
|
| [362] |
Luo SC, Wei SM, Luo XT, et al. (2024) How probiotics, prebiotics, synbiotics, and postbiotics prevent dental caries: An oral microbiota perspective. NP J Biofilms Microbiomes 10: 14. https://doi.org/10.1038/s41522-024-00488-7
|
| [363] |
Suresh N, Joseph B, Sathyan P, et al. (2024) Photodynamic therapy: An emerging therapeutic modality in dentistry. Bioorg Med Chem 114: 117962. https://doi.org/10.1016/j.bmc.2024.117962
|
| [364] |
Wright PP, Ramachandra SS (2022) Quorum sensing and quorum quenching with a focus on cariogenic and periodontopathic oral biofilms. Microorganisms 10: 1783. https://doi.org/10.3390/microorganisms10091783
|
| [365] |
Guo X, Wang X, Shi J, et al. (2024) A review and new perspective on oral bacteriophages: Manifestations in the ecology of oral diseases. J Oral Microbiol 16: 2344272. https://doi.org/10.1080/20002297.2024.2344272
|
| [366] |
Maitre Y, Mahalli R, Micheneau P, et al. (2021) Pre and probiotics involved in the modulation of oral bacterial species: New therapeutic leads in mental disorders?. Microorganisms 9: 1450. https://doi.org/10.3390/microorganisms9071450
|
| [367] | Borrego-Ruiz A, Borrego JJ (2024) Pychobiotics: A new perspective on the treatment of stress, anxiety, and depression. Anxiety Stress 30: 79-93. https://doi.org/10.5093/anyes2024a11 |
| [368] |
Borrego-Ruiz A, Borrego JJ (2024) Microbial dysbiosis in the skin microbiome and its psychological consequences. Microorganisms 12: 1908. https://doi.org/10.3390/microorganisms12091908
|
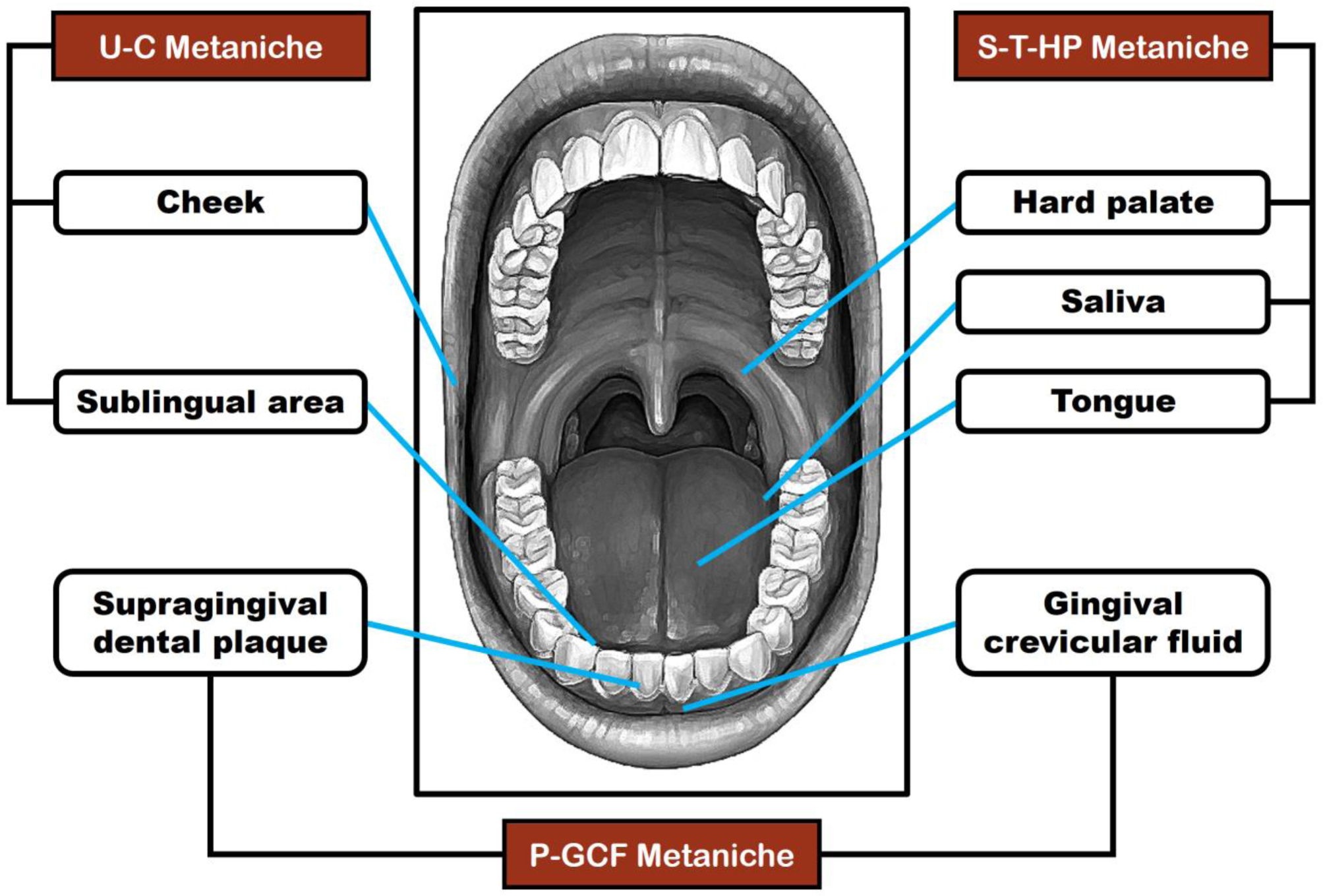
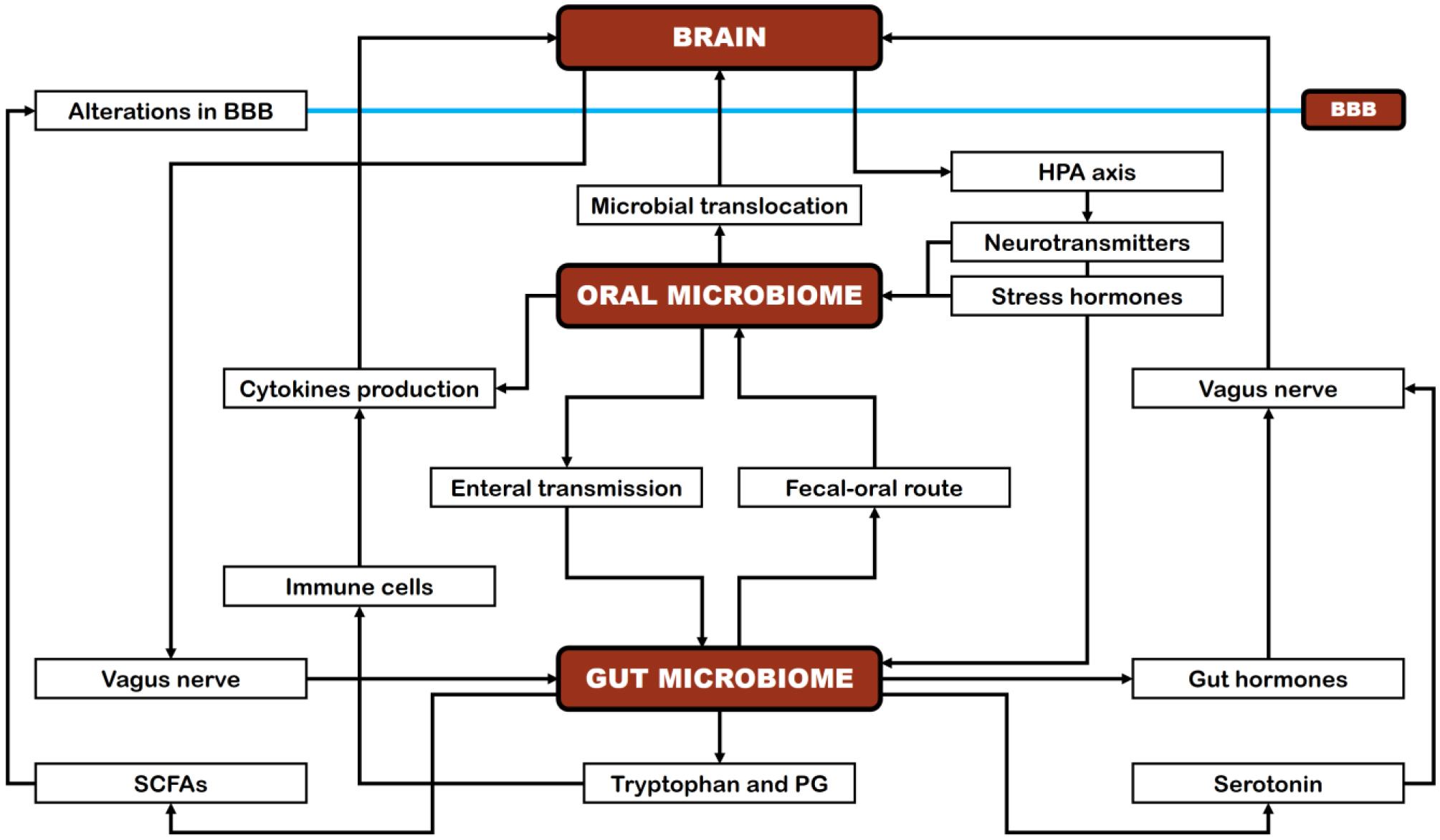
Figures(2) / Tables(2)

Alejandro Borrego-Ruiz, Juan J. Borrego. Human oral microbiome and its influence on mental health and brain disorders[J]. AIMS Microbiology, 2025, 11(2): 242-294. doi: 10.3934/microbiol.2025013







 DownLoad:
DownLoad: