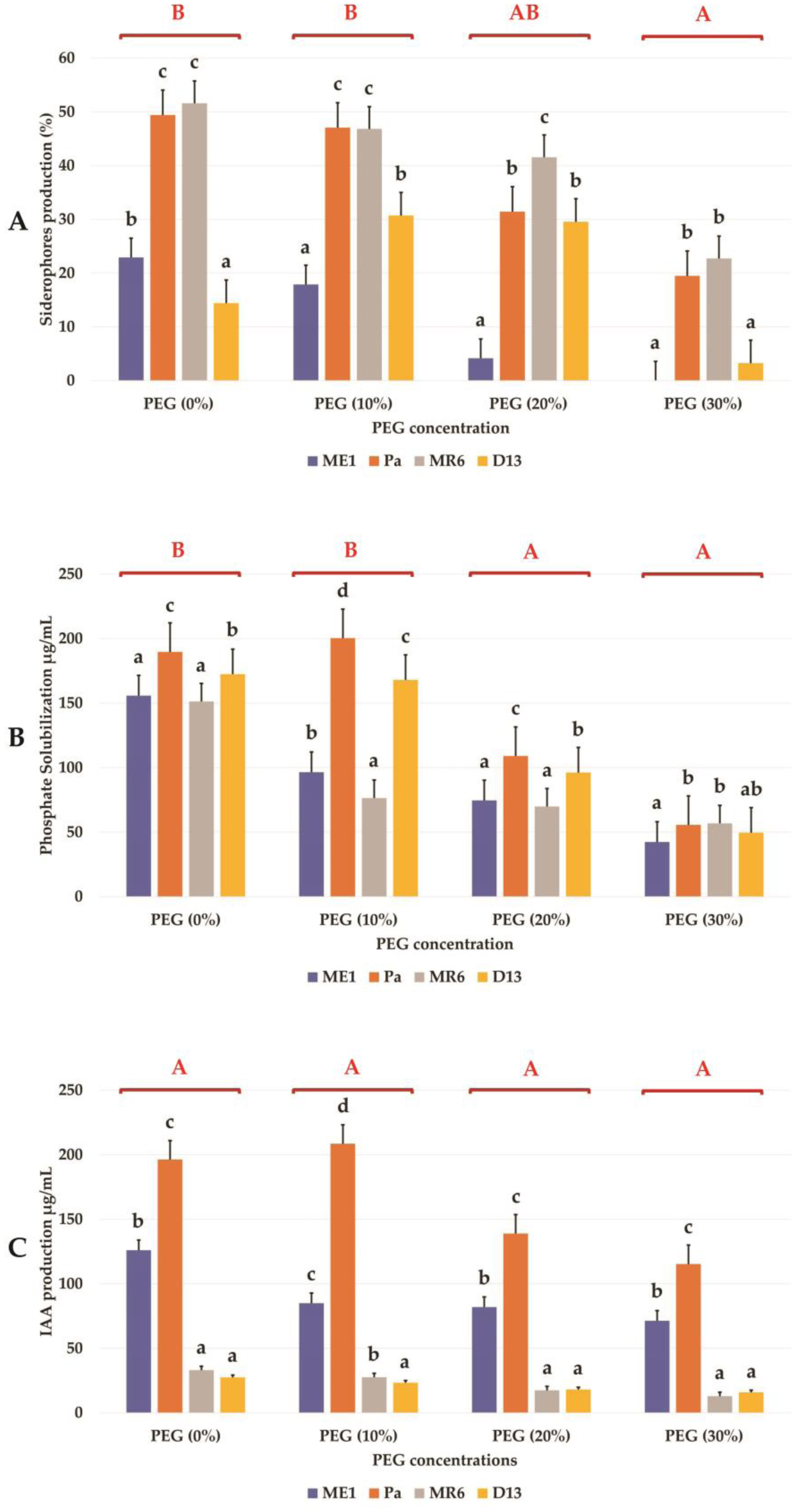
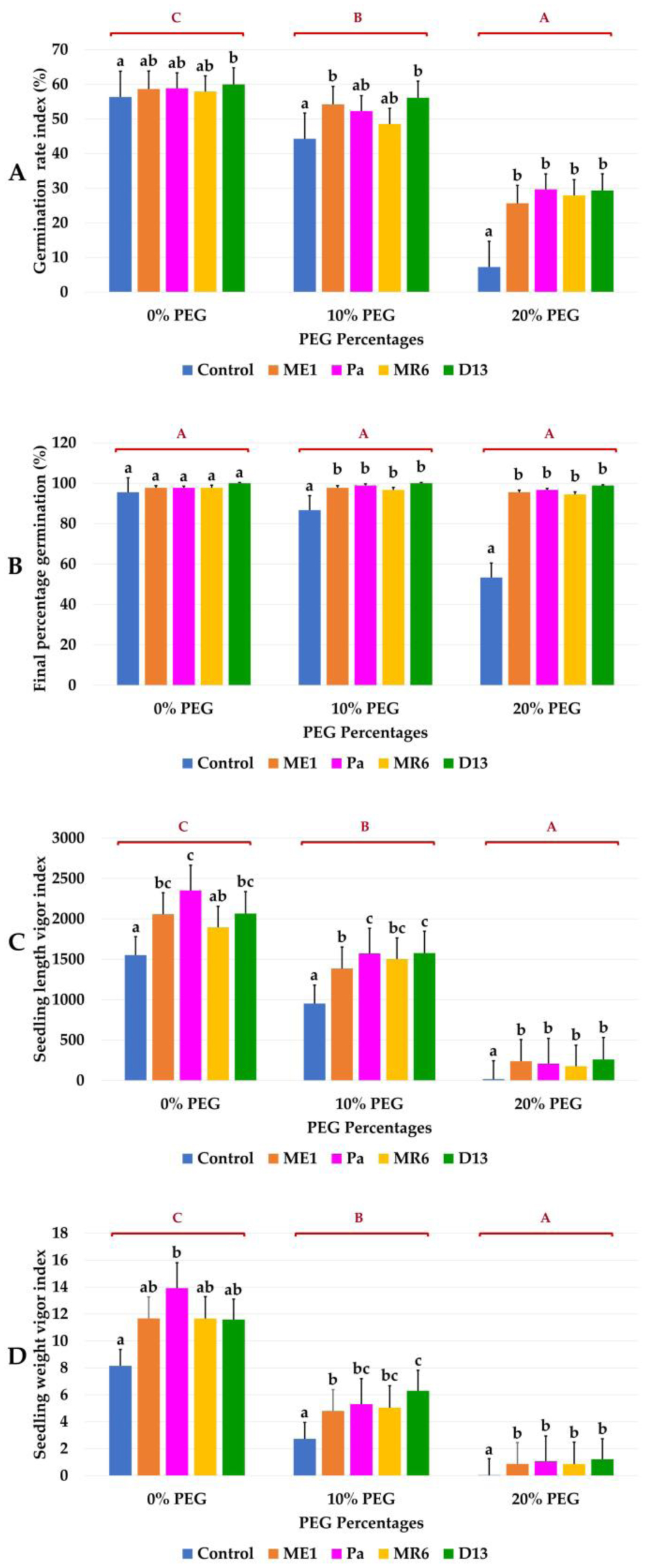
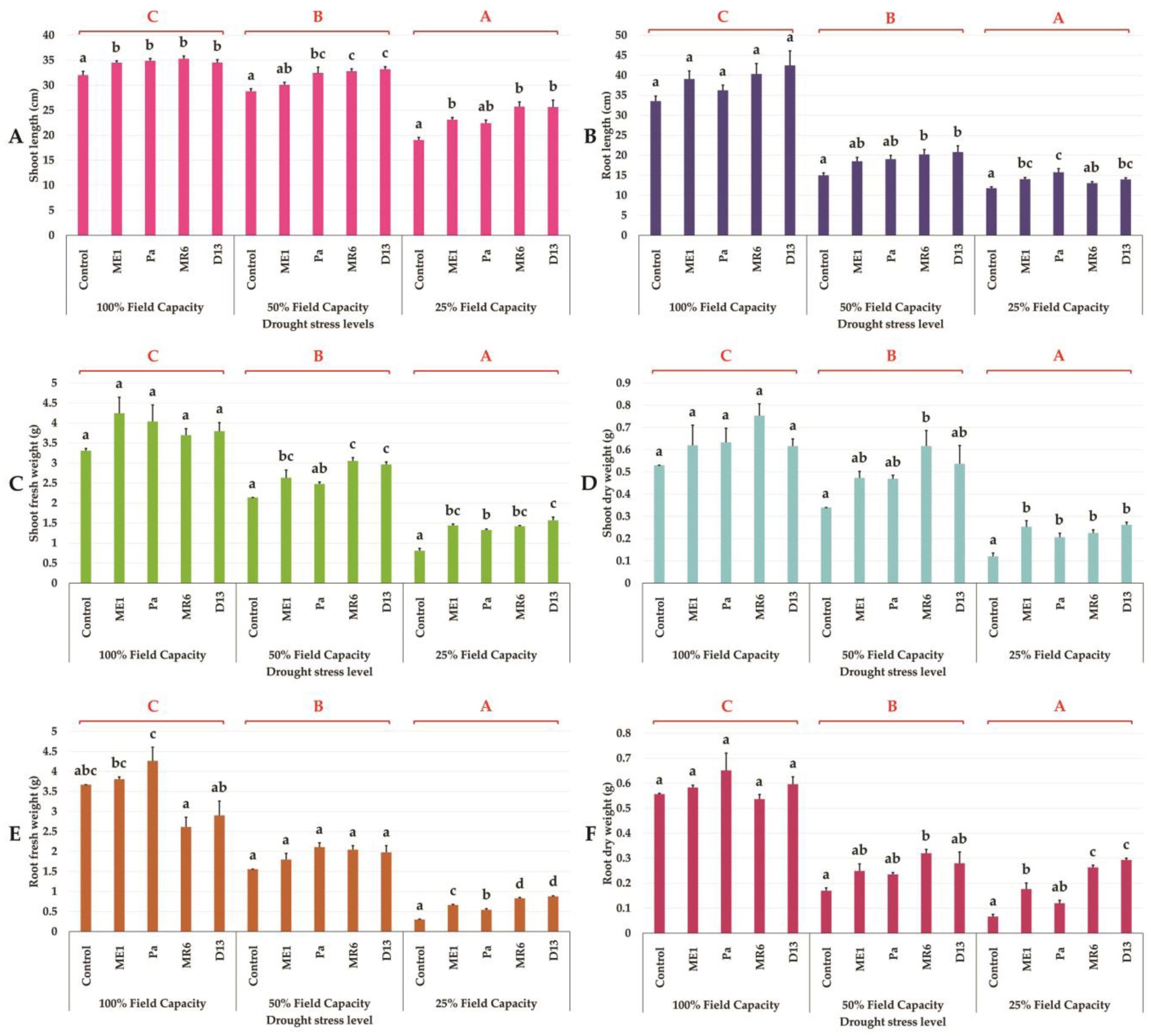
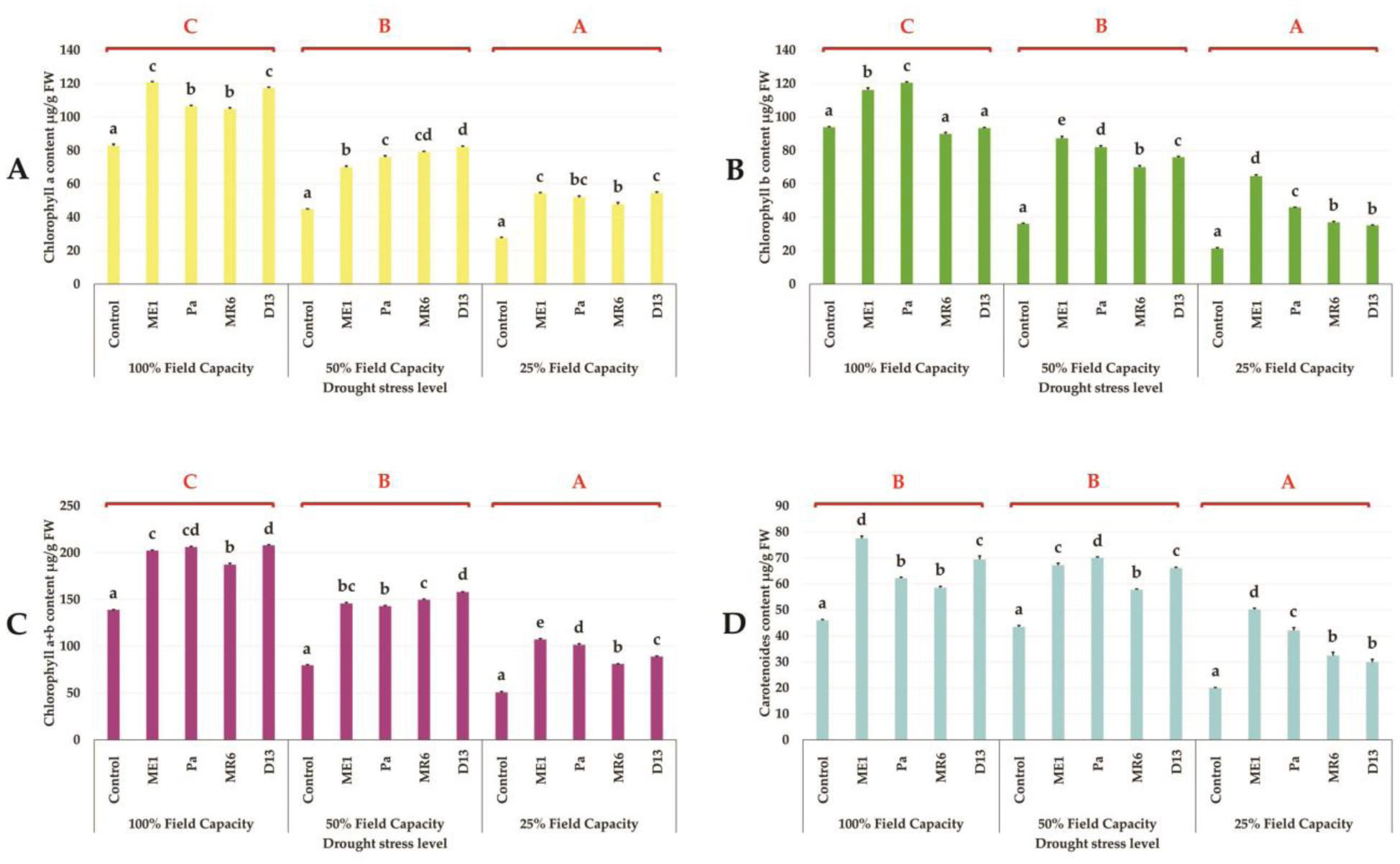
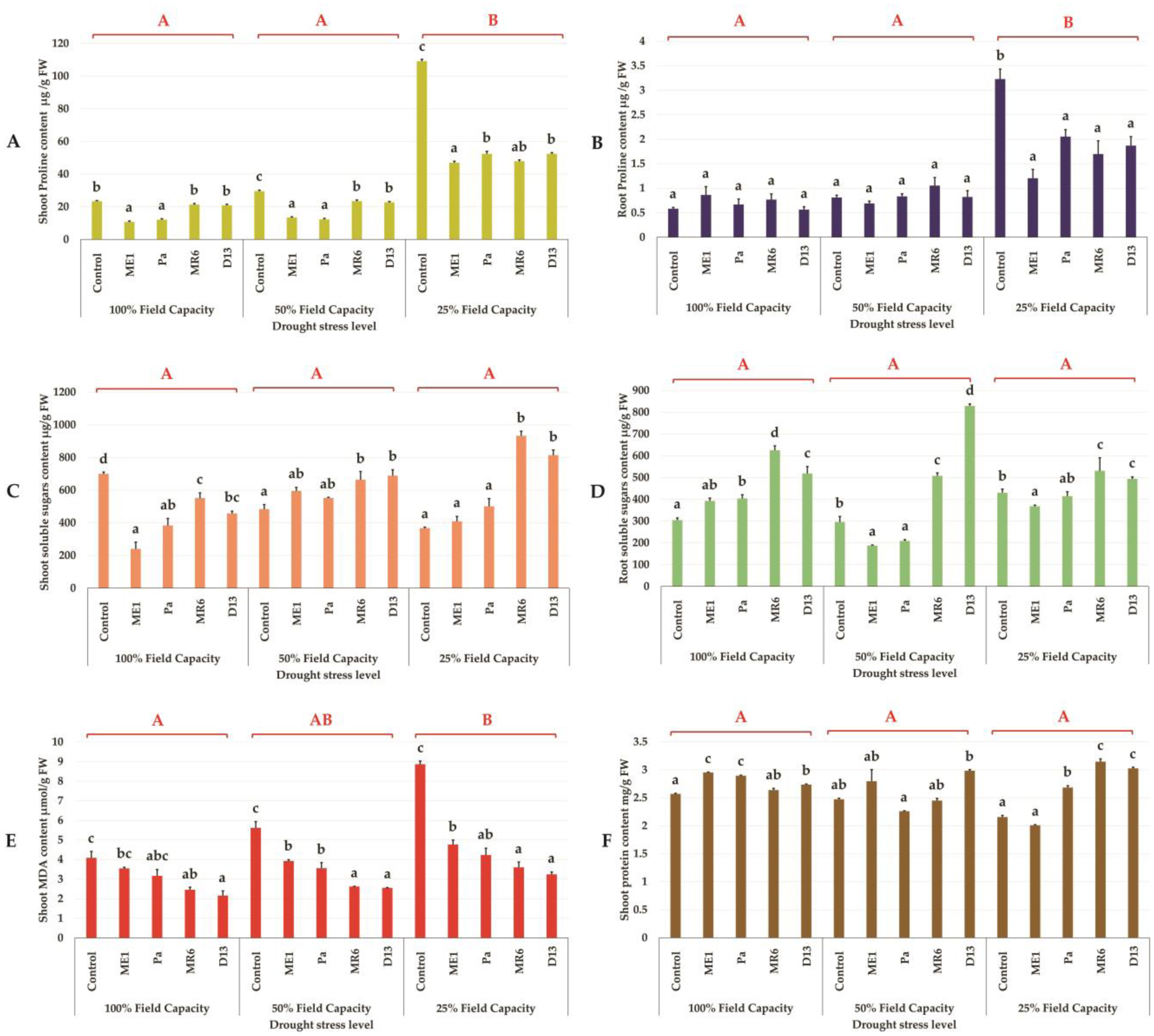
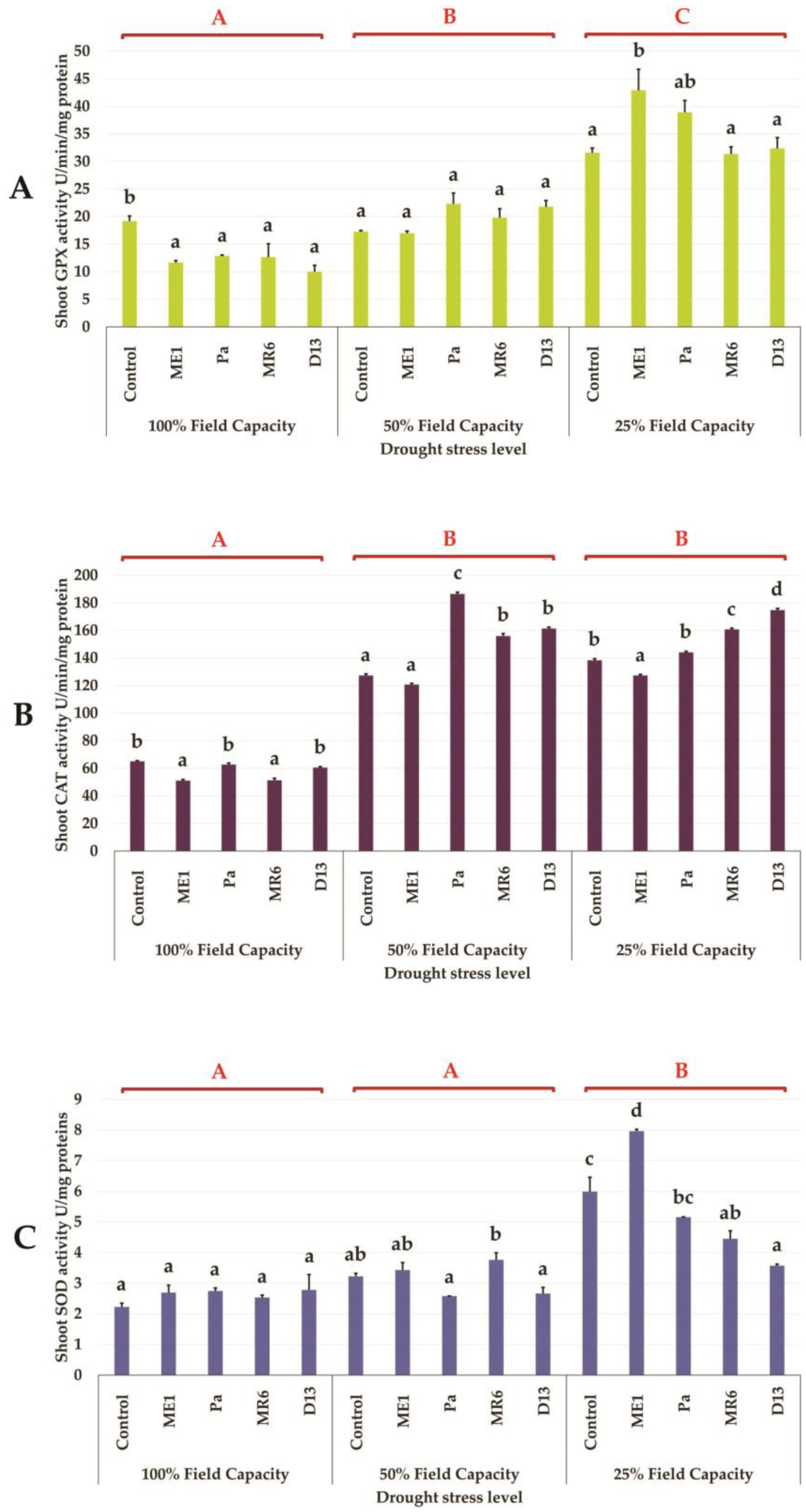
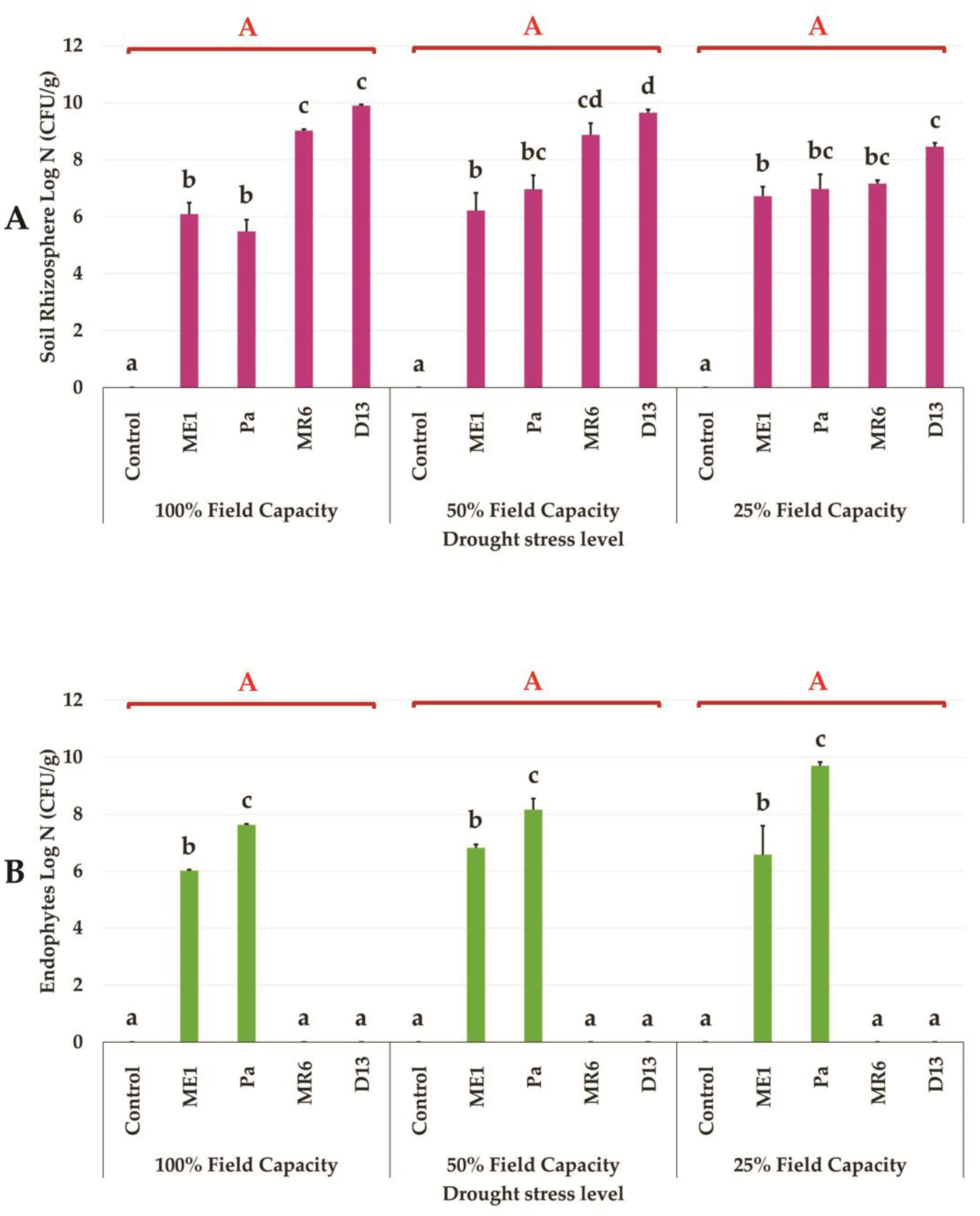
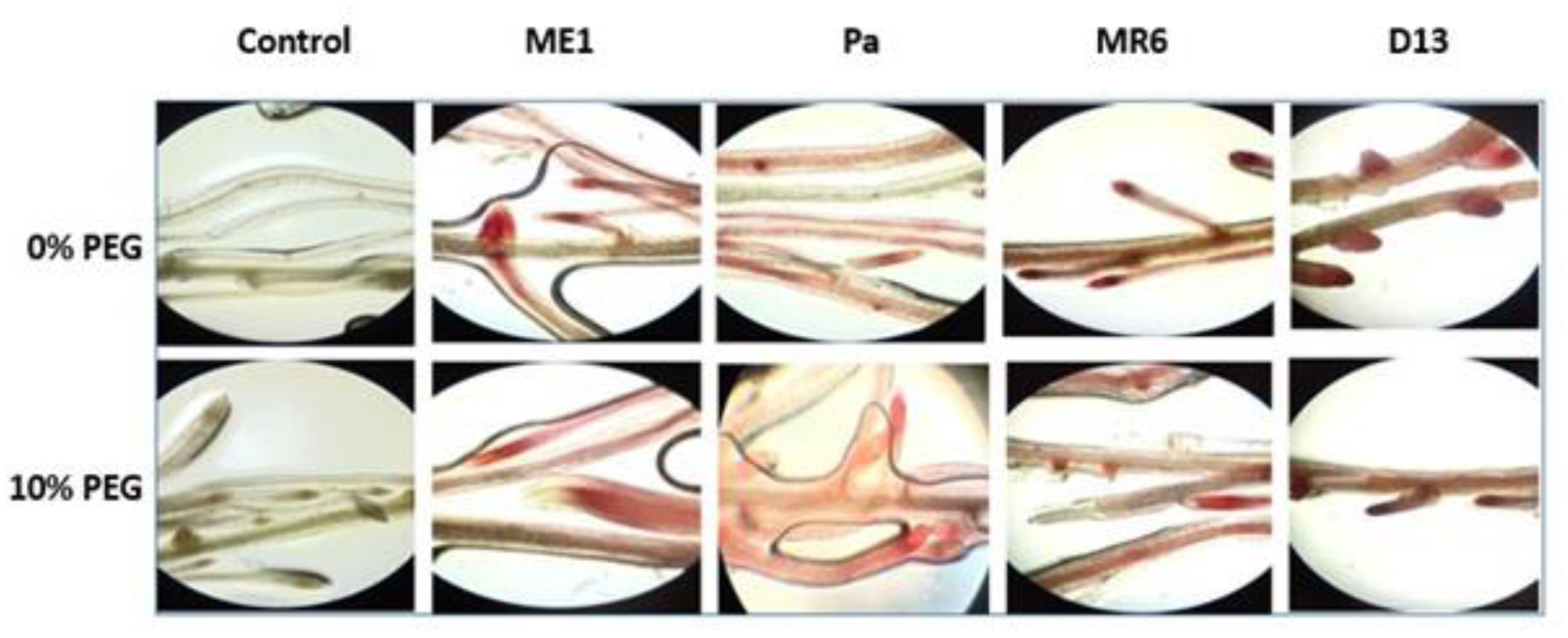
Drought stress represents a major constraint with significant impacts on wheat crop globally. The use of plant growth-promoting bacteria (PGPB) has emerged as a promising strategy to alleviate the detrimental impacts of water stress and enhance plant development. We investigated 24 strains from diverse ecosystems, assessed for PGP traits and tolerance ability to abiotic stresses (drought, salinity, temperature, pH, heavy metals, pollutants, herbicides, and fungicides). The most effective bacterial strains Providencia vermicola ME1, Pantoea agglomerans Pa, Pseudomonas knackmussi MR6, and Bacillus sp D13 were chosen. Furthermore, these strains exhibited PGP activities under osmotic stress (0, 10, 20, and 30% PEG-6000). The impact of these osmotolerant PGPBs on wheat (Triticum durum L.) growth under drought stress was assessed at two plant growth stages. In an in vitro wheat seed germination experiment, bacterial inoculation significantly enhanced germination parameters. In pot experiments, the potential of these bacteria was evaluated in wheat plants under three treatments: Well-watered (100% field capacity), moderate stress (50% FC), and severe stress (25% FC). Results showed a significant decline in wheat growth parameters under increasing water stress for uninoculated seedlings. In contrast, bacterial inoculation mitigated these adverse effects, significantly improving morphological parameters and chlorophyll pigment contents under the stress conditions. While malondialdehyde (lipid peroxidation) and proline contents increased significantly with drought intensity, they decreased after bacterial inoculation. The antioxidant enzyme activities (GPX, CAT, and SOD) in plants decreased after bacterial inoculation. The increased root colonization capacity observed under water stress was attributed to their ability to favorable adaptations in a stressful environment. This study highlighted the potential of selected PGPB to alleviate water stress effects on wheat, promoting practical applications aimed at enhancing crop resilience under conditions of water shortage.
Citation: Naoual Bouremani, Hafsa Cherif-Silini, Allaoua Silini, Nour El Houda Rabhi, Ali Chenari Bouket, Lassaad Belbahri. Osmotolerant plant growth promoting bacteria mitigate adverse effects of drought stress on wheat growth[J]. AIMS Microbiology, 2024, 10(3): 507-541. doi: 10.3934/microbiol.2024025
Drought stress represents a major constraint with significant impacts on wheat crop globally. The use of plant growth-promoting bacteria (PGPB) has emerged as a promising strategy to alleviate the detrimental impacts of water stress and enhance plant development. We investigated 24 strains from diverse ecosystems, assessed for PGP traits and tolerance ability to abiotic stresses (drought, salinity, temperature, pH, heavy metals, pollutants, herbicides, and fungicides). The most effective bacterial strains Providencia vermicola ME1, Pantoea agglomerans Pa, Pseudomonas knackmussi MR6, and Bacillus sp D13 were chosen. Furthermore, these strains exhibited PGP activities under osmotic stress (0, 10, 20, and 30% PEG-6000). The impact of these osmotolerant PGPBs on wheat (Triticum durum L.) growth under drought stress was assessed at two plant growth stages. In an in vitro wheat seed germination experiment, bacterial inoculation significantly enhanced germination parameters. In pot experiments, the potential of these bacteria was evaluated in wheat plants under three treatments: Well-watered (100% field capacity), moderate stress (50% FC), and severe stress (25% FC). Results showed a significant decline in wheat growth parameters under increasing water stress for uninoculated seedlings. In contrast, bacterial inoculation mitigated these adverse effects, significantly improving morphological parameters and chlorophyll pigment contents under the stress conditions. While malondialdehyde (lipid peroxidation) and proline contents increased significantly with drought intensity, they decreased after bacterial inoculation. The antioxidant enzyme activities (GPX, CAT, and SOD) in plants decreased after bacterial inoculation. The increased root colonization capacity observed under water stress was attributed to their ability to favorable adaptations in a stressful environment. This study highlighted the potential of selected PGPB to alleviate water stress effects on wheat, promoting practical applications aimed at enhancing crop resilience under conditions of water shortage.

| [1] | Gowtham HG, Singh SB, Shilpa N, et al. (2022) Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants 11: 1763. https://doi.org/10.3390/antiox11091763 |
| [2] | Hossain A, Hassan Z, Sohag MH, et al. (2023) Impact of the endophytic and rhizospheric bacteria on crop development: prospects for advancing climate-smart agriculture. J Crop Sci Biotechnol 26: 405-431. https://doi.org/10.1007/s12892-023-00195-3 |
| [3] | Papadopoulou A, Theodora M, Nathalie K, et al. (2022) Decoding the potential of a new Pseudomonas Putida strain for inducing drought tolerance of tomato (Solanum Lycopersicum) plants through seed biopriming. J Plant Physiol 271: 153658. https://doi.org/10.1016/j.jplph.2022.153658 |
| [4] | Chieb M, Gachomo EW (2023) The role of plant growth promoting rhizobacteria in plant drought stress responses. BMC Plant Biol 23: 407. https://doi.org/10.1186/s12870-023-04403-8 |
| [5] | Romero-Munar A, Aroca R, Zamarreño AM, et al. (2023) Dual inoculation with Rhizophagus irregularis and Bacillus megaterium improves maize tolerance to combined drought and high temperature stress by enhancing root hydraulics, photosynthesis and hormonal responses. Int J Mol Sci 24: 5193. https://doi.org/10.1186/s12870-023-04403-8 |
| [6] | Shirmohammadi E, Alikhani HA, Pourbabaei AA, et al. (2020) Improved phosphorus (p) uptake and yield of rainfed wheat fed with p fertilizer by drought-tolerant phosphate-solubilizing fluorescent pseudomonads strains: A field study in drylands. J Soil Sci Plant Nutr 20: 2195-2211. https://doi.org/10.1007/s42729-020-00287-x |
| [7] | Drought and agricultureFood and Agriculture Organization of the United Nations. Available from: https://www.fao.org/land-water/water/drought/droughtandag/en/ |
| [8] | Gao X, Luan J, Wang L, et al. (2023) Effect of the plant growth promoting rhizobacterium, Cronobacter sp. Y501, for enhancing drought tolerance in maize (Zea mays L.). J Soil Sci Plant Nutr 23: 2786-2797. https://doi.org/10.1007/s42729-023-01234-2 |
| [9] | Arora S, Jha PN (2023) Drought-tolerant Enterobacter bugandensis WRS7 induces systemic tolerance in Triticum aestivum L. (wheat) under drought conditions. J Plant Growth Regul 42: 7715-7730. https://doi.org/10.1007/s00344-023-11044-6 |
| [10] | Kour D, Yadav AN (2022) Bacterial mitigation of drought stress in plants: Current perspectives and future challenges. Curr Microbiol 79: 248. https://doi.org/10.1007/s00284-022-02939-w |
| [11] | Jha Y, Yadav KA, Mohamed HI (2023) Plant growth-promoting bacteria and exogenous phytohormones alleviate the adverse effects of drought stress in pigeon pea plants. Plant Soil . https://doi.org/10.1007/s11104-023-06155-8 |
| [12] | Akhtar N, Ilyas N, Mashwani Z, et al. (2021) Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nano-particles for amelioration of drought stress in wheat. Plant Physiol Biochem 166: 160-176. https://doi.org/10.1016/j.plaphy.2021.05.039 |
| [13] | Valizadeh-rad K, Motesharezadeh B, Alikhani HA, et al. (2023) Morphophysiological and nutritional responses of canola and wheat to water deficit stress by the application of plant growth-promoting bacteria, nano-silicon, and silicon. J Plant Growth Regul 42: 3615-3631. https://doi.org/10.1007/s00344-022-10824-w |
| [14] | Nikhil PT, Faiz U, Mohapatra S (2023) The drought-tolerant rhizobacterium, Pseudomonas putida AKMP7, suppresses polyamine accumulation under well-watered conditions and diverts putrescine into GABA under water-stress, in Oryza sativa. Enviro Exp Bot 211: 105377. https://doi.org/10.1016/j.envexpbot.2023.105377 |
| [15] | Joshi B, Chaudhary A, Singh H, et al. (2020) Prospective evaluation of individual and consortia plant growth promoting rhizobacteria for drought stress amelioration in rice (Oryza sativa L.). Plant Soil 457: 225-240. https://doi.org/10.1007/s11104-020-04730-x |
| [16] | Gul F, Khan IU, Rutherford S, et al. (2023) Plant growth promoting rhizobacteria and biochar production from Parthenium hysterophorus enhance seed germination and productivity in barley under drought stress. Front Plant Sci 14: 1175097. https://doi.org/10.3389/fpls.2023.1175097 |
| [17] | Umapathi M, Chandrasekhar CN, Senthil A, et al. (2022) Isolation, characterization and plant growth-promoting effects of sorghum [Sorghum bicolor (L.) Moench.] root-associated rhizobacteria and their potential role in drought mitigation. Arch Microbiol 204: 354. https://doi.org/10.1007/s00203-022-02939-1 |
| [18] | Yaghoubian I, Modarres-Sanavy SAM, Smith DL (2022) Plant growth promoting microorganisms (PGPM) as an eco-friendly option to mitigate water deficit in soybean (Glycine max L.): Growth, physio-biochemical properties and oil content. Plant Physiol Biochem 191: 55-66. https://doi.org/10.1016/j.plaphy.2022.09.013 |
| [19] | Ansari FA, Jabeen M, Ahmad I (2021) Pseudomonas azotoformans FAP5, a novel biofilm-forming PGPR strain, alleviates drought stress in wheat plant. Int J Environ Sci Technol 18: 3855-3870. https://doi.org/10.1007/s13762-020-03045-9 |
| [20] | Singh D, Thapa S, Yadav J, et al. (2023) Deciphering the mechanisms of microbe mediated drought stress alleviation in wheat. Acta Physiol Plant 45: 81. https://doi.org/10.1007/s11738-023-03562-3 |
| [21] | Ilyas N, Mumtaz K, Akhtar N, et al. (2020) Exopolysaccharides producing bacteria for the amelioration of drought stress in wheat. Sustainability 12: 8876. https://doi.org/10.3390/su12218876 |
| [22] | Yasmin H, Rashid U, Hassan MN, et al. (2021) Volatile organic compounds produced by Pseudomonas pseudoalcaligenes alleviated drought stress by modulating defense system in maize (Zea mays L.). Physiol Plant 172: 896-911. https://doi.org/10.1111/ppl.13304 |
| [23] | Cherif-Silini H, Silini A, Yahiaoui B, et al. (2016) Phylogenetic and plant-growth-promoting characteristics of Bacillus isolated from the wheat rhizosphere. Ann Microbiol 66: 1087-1097. https://doi.org/10.1007/s13213-016-1194-6 |
| [24] | Kerbab S, Silini A, Bouket AC, et al. (2021) Mitigation of NaCl Stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl Sci 11: 1034. https://doi.org/10.3390/app11031034 |
| [25] | Balla A, Silini A, Cherif-Silini H, et al. (2022) Screening of cellulolytic bacteria from various ecosystems and their cellulases production under multi-stress conditions. Catalysts 12: 769. https://doi.org/10.3390/catal12070769 |
| [26] | Boulahouat S, Cherif-Silini H, Silini A, et al. (2022) Critical Evaluation of biocontrol ability of bayoud infected date palm phyllospheric Bacillus spp. suggests that in vitro selection does not guarantee success in planta. Agronomy 12: 2403. https://doi.org/10.3390/agronomy12102403 |
| [27] | Cherif-Silini H, Thissera B, Bouket AC, et al. (2019) Durum wheat stress tolerance induced by endophyte Pantoea agglomerans with genes contributing to plant functions and secondary metabolite arsenal. Int J Mol Sci 20: 3989. https://doi.org/10.3390/ijms20163989 |
| [28] | Rabhi NEH, Cherif-Silini H, Silini A, et al. (2019) Alleviation of salt stress via habitat-adapted symbiosis. Forests 13: 586. https://doi.org/10.3390/f13040586 |
| [29] | Borriss R, Chen XH, Rueckert C, et al. (2011) Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7T and FZB42T: A proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int J Syst Evol Microbiol 61: 1786-1801. https://doi.org/10.1099/ijs.0.023267-0 |
| [30] | Olsen SR, Sommers LE (1982) Phosphorus. Methods of Soil Analysis Part 2 Chemical and Microbiological Properties . Madison: American Society of Agronomy, Soil Science Society of America 403-430. https://doi.org/10.2134/agronmonogr9.2.2ed.c24 |
| [31] | Schwyn B, Neilands JB (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160: 47-56. https://doi.org/10.1016/0003-2697(87)90612-9 |
| [32] | Slama HB, Triki MA, Bouket AC, et al. (2019) Screening of the high-rhizosphere competent Limoniastrum monopetalum' culturable endophyte microbiota allows the recovery of multifaceted and versatile biocontrol agents. Microorganisms 7: 249. https://doi.org/10.3390/microorganisms7080249 |
| [33] | Dworkin M, Foster J (1958) Experiments with some microorganisms which utilize ethane and hydrogen. J Bacteriol 75: 592-603. https://doi.org/10.1128/jb.75.5.592-603.1958 |
| [34] | Li Z, Chang S, Lin L, et al. (2011) A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Lett Appl Microbiol 53: 178-185. https://doi.org/10.1111/j.1472-765X.2011.03088.x |
| [35] | Zeriouh H, de Vicente A, Pérez-García A, et al. (2014) Surfactin triggers biofilm formation of Bacillus subtilis in melon phylloplane and contributes to the biocontrol activity. Environ Microbiol 16: 2196-2211. https://doi.org/10.1111/1462-2920.12271 |
| [36] | Li X, Peng D, Zhang Y, et al. (2020) Klebsiella sp. PD3, a phenanthrene (PHE)-degrading strain with plant growth promoting properties enhances the PHE degradation and stress tolerance in rice plants. Ecotoxicol Environ Saf 201: 110804. https://doi.org/10.1016/j.ecoenv.2020.110804 |
| [37] | Saadaoui N, Silini A, Cherif-Silini H, et al. (2022) Semi-arid-habitat-adapted plant-growth-promoting rhizobacteria allows efficient wheat growth promotion. Agronomy 12: 2221. https://doi.org/10.3390/agronomy12092221 |
| [38] | Dubois M, Gilles KA, Hamilton JK, et al. (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28: 350-356. https://doi.org/10.1021/ac60111a017 |
| [39] | Lowry OH, Rosbrough NJ, Farr AL, et al. (1951) Protein measurement with the folin phenol reagent. J Bio Chem 193: 265-275. https://doi.org/10.1016/S0021-9258(19)52451-6 |
| [40] | Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473-497. https://doi.org/10.1111/j.1399-3054.1962.tb08052.x |
| [41] | Cherif-Silini H, Silini A, Bouket AC, et al. (2021) Tailoring next generation plant growth promoting microorganisms as versatile tools beyond soil desalinization: A road map towards field application. Sustainability 13: 4422. https://doi.org/10.3390/su13084422 |
| [42] | Rabhi NEH, Silini A, Cherif-Silini H, et al. (2018) Pseudomonas knackmussii MLR6, a rhizospheric strain isolated from halophyte, enhances salt tolerance in Arabidopsis thaliana. J Appl Microbiol 125: 1836-1851. https://doi.org/10.1111/jam.14082 |
| [43] | Noha MA, Bothaina AA, Shereen AM, et al. (2022) Utilization of drought-tolerant bacterial strains isolated from harsh soils as a plant growth-promoting rhizobacteria (PGPR). Saudi J Biol Sci 29: 1760-1769. https://doi.org/10.1016/j.sjbs.2021.10.054 |
| [44] | Al-Shwaiman HA, Shahid M, Elgorban AM, et al. (2022) Beijerinckia fluminensis BFC-33, a novel multi-stress-tolerant soil bacterium: Deciphering the stress amelioration, phytopathogenic inhibition and growth promotion in Triticum aestivum (L.). Chemosphere 295: 133843. https://doi.org/10.1016/j.chemosphere.2022.133843 |
| [45] | Silambarasan S, Logeswari P, Vangnai AS, et al. (2022) Plant growth-promoting actinobacterial inoculant assisted phytoremediation increases cadmium uptake in Sorghum bicolor under drought and heat stresses. Environ Pollut 307: 119489. https://doi.org/10.1016/j.envpol.2022.11948 |
| [46] | Khan A, Singh AV (2021) Multifarious effect of ACC deaminase and EPS producing Pseudomonas sp. and Serratia marcescens to augment drought stress tolerance and nutrient status of wheat. World J Microbiol Biotechnol 37: 198. https://doi.org/10.1007/s11274-021-03166-4 |
| [47] | Latif M, Bukhari SAH, Alrajhi AA, et al. (2022) Inducing drought tolerance in wheat through exopolysaccharide-producing rhizobacteria. Agronomy 12: 1140. https://doi.org/10.3390/agronomy12051140 |
| [48] | Gowtham HG, Brijesh SS, Murali M, et al. (2020) Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiol Res 234: 126422. https://doi.org/10.1016/j.micres.2020.126422 |
| [49] | Rashid U, Yasmin H, Hassan MN, et al. (2022) Drought-tolerant Bacillus megaterium isolated from semi-arid conditions induces systemic tolerance of wheat under drought conditions. Plant Cell Rep 41: 549-569. https://doi.org/10.1007/s00299-020-02640-x |
| [50] | Zhao T, Deng X, Xiao Q, et al. (2020) IAA priming improves the germination and seedling growth in cotton (Gossypium hirsutum L.) via regulating the endogenous phytohormones and enhancing the sucrose metabolism. Ind Crops Prod 155: 112788. https://doi.org/10.1016/j.indcrop.2020.112788 |
| [51] | Kasim WA, Osman MEH, Omar MN, et al. (2021) Enhancement of drought tolerance in Triticum aestivum L. seedlings using Azospirillum brasilense NO40 and Stenotrophomonas maltophilia B11. Bull Natl Res Cent 45: 95. https://doi.org/10.1186/s42269-021-00546-6 |
| [52] | Batool T, Ali S, Seleiman MF, et al. (2020) Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci Rep 10: 16975. https://doi.org/10.1038/s41598-020-73489-z |
| [53] | Khalilpour M, Mozafari V, Abbaszadeh-Dahaji P (2021) Tolerance to salinity and drought stresses in pistachio (Pistacia vera L.) seedlings inoculated with indigenous stress-tolerant PGPR isolates. Sci Hort 289: 110440. https://doi.org/10.1016/j.scienta.2021.110440 |
| [54] | Murali M, Singh SB, Gowtham HG, et al. (2021) Induction of drought tolerance in Pennisetum glaucum by ACC deaminase producing PGPR-Bacillus amyloliquefaciens through Antioxidant defense system. Microbiol Res 253: 126891. https://doi.org/10.1016/j.micres.2021.126891 |
| [55] | Ma Y, Rajkumar M, Moreno A, et al. (2017) Serpentine endophytic bacterium Pseudomonas azotoformans ASS1 accelerates phytoremediation of soil metals under drought stress. Chemosphere 185: 75-85. https://doi.org/10.1016/j.chemosphere.2017.06.135 |
| [56] | Zarei T, Moradi A, Kazemeini SA, et al. (2020) The role of ACC deaminase producing bacteria in improving sweet corn (Zea mays L. var. Saccharata) productivity under limited availability of irrigation water. Sci Rep 10: 20361. https://doi.org/10.1038/s41598-020-77305-6 |
| [57] | Khan N, Bano A (2019) Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS One 14: 0222302. https://doi.org/10.1371/journal.pone.0222302 |
| [58] | Sheteiwy MS, Abd Elgawad H, Xiong YC, et al. (2021) Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol Plant 172: 2153-2169. https://doi.org/10.1111/ppl.13454 |
| [59] | Sood G, Kaushal R, Sharma M (2020) Significance of inoculation with Bacillus subtilis to alleviate drought stress in wheat (Triticum aestivum L.). Vegetos 33: 782-792. https://doi.org/10.1007/s42535-020-00149-y |
| [60] | Han L, Zhang M, Du L, et al. (2022) Effects of Bacillus amyloliquefaciens QST713 on photosynthesis and antioxidant characteristics of alfalfa (Medicago sativa L.) under drought stress. Agronomy 12: 2177. https://doi.org/10.3390/agronomy12092177 |
| [61] | Niu X, Song L, Xiao Y, et al. (2018) Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front Microbiol 8: 2580. https://doi.org/10.3389/fmicb.2017.02580 |
| [62] | Rashid U, Yasmin H, Hassan M N, et al. (2022) Drought-tolerant Bacillus megaterium isolated from semi-arid conditions induces systemic tolerance of wheat under drought conditions. Plant Cell Rep 41: 549-569. https://doi.org/10.1007/s00299-020-02640-x |
| [63] | Singh RP, Jha PN (2017) The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front Microbiol 8: 1945. https://doi.org/10.3389/fmicb.2017.01945 |
 microbiol-10-03-025-s001.pdf microbiol-10-03-025-s001.pdf |
 |


Figures(10) / Tables(1)

Naoual Bouremani, Hafsa Cherif-Silini, Allaoua Silini, Nour El Houda Rabhi, Ali Chenari Bouket, Lassaad Belbahri. Osmotolerant plant growth promoting bacteria mitigate adverse effects of drought stress on wheat growth[J]. AIMS Microbiology, 2024, 10(3): 507-541. doi: 10.3934/microbiol.2024025







 DownLoad:
DownLoad: