Citation: Alexey V. Rakov, Natalya A. Kuznetsova, Anatoly A. Yakovlev. Genetic diversity of Salmonella enterica subsp. enterica serovar Enteritidis in the Siberia and Far East of Russia based on plasmid profiles[J]. AIMS Microbiology, 2020, 6(2): 106-120. doi: 10.3934/microbiol.2020007

| [1] | Ryan MP, O'Dwyer J, Adley CC (2017) Evaluation of the complex nomenclature of the clinically and veterinary significant pathogen Salmonella. BioMed Res Int 2017: 3782182. |
| [2] |
Lamas A, Miranda JM, Regal P, et al. (2018) A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol Res 206: 60-73. doi: 10.1016/j.micres.2017.09.010

|
| [3] | Shubin FN, Rakov AV, Kuznetsova NA, et al. (2015) Zoonotic salmonellosis in the Far East: main aspects of a problem. Far-Eastern J Infect Pathol 29: 72-79. |
| [4] | EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control) (2018) The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J 16: 5500. |
| [5] |
Hendriksen RS, Vieira AR, Karlsmose S, et al. (2011) Global monitoring of Salmonella serovar distribution from the world health organization global foodborne infections network country data bank: results of quality assured laboratories from 2001 to 2007. Foodborne Pathog Dis 8: 887-900. doi: 10.1089/fpd.2010.0787
|
| [6] |
Shubin FN (2015) Zoonotic salmonellosis in Russia: main aspects of a problem. Epidemiol Vaccinal Prev 80: 28-30. doi: 10.31631/2073-3046-2015-14-1-28-30
|
| [7] |
Rakov AV, Mastriani E, Liu SL, et al. (2019) Association of Salmonella virulence factor alleles with intestinal and invasive serovars. BMC Genomics 20: 429. doi: 10.1186/s12864-019-5809-8
|
| [8] |
Dagan T, Blekhman R, Graur D (2006) The ‘domino theory’ of gene death: gradual and mass gene extinction events in three lineages of obligate symbiotic bacterial pathogens. Mol Biol Evol 23: 310-316. doi: 10.1093/molbev/msj036
|
| [9] |
Nuccio SP, Bäumler AJ (2014) Comparative analysis of Salmonella genomes identifies a metabolic network for escalating growth in the inflamed gut. mBio 5: e00929-14. doi: 10.1128/mBio.00929-14
|
| [10] |
Valenzuela LM, Hidalgo AA, Rodríguez L, et al. (2015) Pseudogenization of sopA and sopE2 is functionally linked and contributes to virulence of Salmonella enterica serovar Typhi. Infect Genet Evol 33: 131-142. doi: 10.1016/j.meegid.2015.04.021
|
| [11] |
Ortega AP, Villagra NA, Urrutia IM, et al. (2016) Lose to win: marT pseudogenization in Salmonella enterica serovar Typhi contributed to the surV-dependent survival to H2O2, and inside human macrophage-like cells. Infect Genet Evol 45: 111-121. doi: 10.1016/j.meegid.2016.08.029
|
| [12] |
Call DR, Kang MS, Daniels J, et al. (2006) Assessing genetic diversity in plasmids from Escherichia coli and Salmonella enterica using a mixed-plasmid microarray. J Appl Microbiol 100: 15-28. doi: 10.1111/j.1365-2672.2005.02775.x
|
| [13] | Shubin FN, Kovalchuk NI, Kuznetsova NA, et al. (2002) Microbiological monitoring for Salmonella enteritidis in Primorye Region. Phenotypical and plasmid characterization of the pathogen. Epidemiol Infect Dis 1: 36-40. |
| [14] |
Rakov AV, Shubin FN (2019) Comparative genomics analysis of Salmonella enterica subsp. enterica serotype Enteritidis virulence plasmid. Russ J Genet 55: 144-153. doi: 10.1134/S102279541902011X
|
| [15] |
Orskov F, Orskov I (1983) Summary of a workshop on the clone concept in the epidemiology, taxonomy and evolution of the Enterobacteriaceae and other bacteria. J Infect Dis 148: 346-357. doi: 10.1093/infdis/148.2.346
|
| [16] | Selander RK, Caugant DA, Whittman TS (1987) Genetic structure and variation in natural populations of Escherichia coli. Cell Mol Biol 2: 1625-1648. |
| [17] |
Kado CI, Liu ST (1981) Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol 145: 1365-1373. doi: 10.1128/JB.145.3.1365-1373.1981
|
| [18] |
Jones GW, Rabert DK, Svinarich DM, et al. (1982) Association of adhesive, invasive, and virulent phenotypes of Salmonella Typhimurium with autonomous 60-megadalton plasmids. Infect Immun 38: 476-486. doi: 10.1128/IAI.38.2.476-486.1982
|
| [19] |
Nakamura M, Sato S, Ohya T, et al. (1985) Possible relationship of a 36-megadalton Salmonella Enteritidis plasmid to virulence in mice. Infect Immun 47: 831-833. doi: 10.1128/IAI.47.3.831-833.1985
|
| [20] |
Rivera MJ, Rivera N, Castillo J, et al. (1991) Molecular and epidemiological study of Salmonella clinical isolates. J Clin Microbiol 29: 927-932. doi: 10.1128/JCM.29.5.927-932.1991
|
| [21] |
Rodrigue DC, Cameron DN, Puhr ND, et al. (1992) Comparison of plasmid profiles, phage types, and antimicrobial resistance patterns of Salmonella Enteritidis isolates in the United States. J Clin Microbiol 30: 854-857. doi: 10.1128/JCM.30.4.854-857.1992
|
| [22] |
Holmberg SD, Wachsmuth IK, Hickman-Brenner FW, et al. (1984) Comparison of plasmid profile analysis, phage typing, and antimicrobial susceptibility testing in characterizing Salmonella Typhimurium isolates from outbreaks. J Clin Microbiol 19: 100-104. doi: 10.1128/JCM.19.2.100-104.1984
|
| [23] |
Farrar WE (1983) Molecular analysis of plasmids in epidemiologic investigation. J Infect Dis 148: 1-6. doi: 10.1093/infdis/148.1.1
|
| [24] |
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98: 503-517. doi: 10.1016/S0022-2836(75)80083-0
|
| [25] |
Olsen JE, Skov MN, Threlfall EJ, et al. (1994) Clonal lines of Salmonella enterica serotype Enteritidis documented by IS200-, ribo-, pulsed-field gel electrophoresis and RFLP typing. J Med Microbiol 40: 15-22. doi: 10.1099/00222615-40-1-15
|
| [26] |
Guerra B, Landeras E, Gonzalez-Hevia MA, et al. (1997) A three-way ribotyping scheme for Salmonella serotype Typhimurium and its usefulness for phylogenetic and epidemiological purposes. J Med Microbiol 46: 307-313. doi: 10.1099/00222615-46-4-307
|
| [27] |
Stanley J, Baquar N, Threlfall EJ (1993) Genotypes and phylogenic relationships of Salmonella Typhimurium are defined by molecular fingerprinting of IS200 and 16S rrn loci. J Gen Microbiol 139: 1133-1140. doi: 10.1099/00221287-139-6-1133
|
| [28] |
Hulton CS, Higgins CF, Sharp PM (1991) ERIC sequences: a novel family of repetitive elements in the genomes of Escherichia coli, Salmonella Typhimurium and other enterobacteria. Mol Microbiol 5: 825-834. doi: 10.1111/j.1365-2958.1991.tb00755.x
|
| [29] | Hilton AC, Banks JG, Penn CW (1996) Random application of polymorphic DNA (RAPD) of Salmonella: strain differentiation and characterization of amplified sequences. J Appl Bacteriol 81: 575-584. |
| [30] |
Schwartz DC, Cantor CR (1984) Separation of yeast chromosome-sized DNAs by pulsed gel electrophoresis. Cell 37: 67-75. doi: 10.1016/0092-8674(84)90301-5
|
| [31] |
Swaminathan B, Barrett TJ, Hunter SB, et al. (2001) PulseNet: the molecular subtyping network for foodborne bacterial disease surveillance, United States. Emerg Infect Dis 7: 382-389. doi: 10.3201/eid0703.017303
|
| [32] |
Ridley AM, Threlfall EJ, Rowe B (1998) Genotypic characterization of Salmonella Enteritidis phage types by plasmid analysis, ribotyping, and pulsed-field gel electrophoresis. J Clin Microbiol 36: 2314-2321. doi: 10.1128/JCM.36.8.2314-2321.1998
|
| [33] |
Ramisse V, Houssu P, Hernandez E, et al. (2004) Variable number of tandem repeats in Salmonella enterica subsp. enterica for typing purposes. J Clin Microbiol 42: 5722-5730. doi: 10.1128/JCM.42.12.5722-5730.2004
|
| [34] |
Boxrud D, Pederson-Gulrud K, Wotton J, et al. (2007) Comparison of multiple-locus variable-number tandem repeat analysis, pulsed-field gel electrophoresis, and phage typing for subtype analysis of Salmonella enterica serotype Enteritidis. J Clin Microbiol 45: 536-543. doi: 10.1128/JCM.01595-06
|
| [35] |
Liu Y, Shi X, Li Y, et al. (2016) The evaluation and application of multilocus variable number tandem repeat analysis (MLVA) for the molecular epidemiological study of Salmonella enterica subsp. enterica serovar Enteritidis infection. Ann Clin Microbiol Antimicrob 15: 4. doi: 10.1186/s12941-016-0119-3
|
| [36] |
Kotetishvili M, Stine OC, Kreger A, et al. (2002) Multilocus sequence typing for characterization of clinical and environmental Salmonella strains. J Clin Microbiol 40: 1626-1635. doi: 10.1128/JCM.40.5.1626-1635.2002
|
| [37] |
Achtman M, Wain J, Weill FX, et al. (2012) Multilocus sequence typing as a replacement for serotyping in Salmonella enterica. PLoS Pathog 8: e1002776. doi: 10.1371/journal.ppat.1002776
|
| [38] |
Feil EJ, Li BC, Aanensen DM, et al. (2004) eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol 186: 1518-1530. doi: 10.1128/JB.186.5.1518-1530.2004
|
| [39] |
Deng X, Shariat N, Driebe EM, et al. (2015) Comparative analysis of subtyping methods against a whole-genome-sequencing standard for Salmonella enterica serotype Enteritidis. J Clin Microbiol 53: 212-218. doi: 10.1128/JCM.02332-14
|
| [40] |
Alikhan NF, Zhou Z, Sergeant MJ, et al. (2018) A genomic overview of the population structure of Salmonella. PLoS Genet 14: e1007261. doi: 10.1371/journal.pgen.1007261
|
| [41] | Rivera MJ, Rivera A, Castillo J, et al. (1993) Plasmid profile in epidemiological studies of human Salmonella infections. J Chemother 5: 288-290. |
| [42] |
Rychlik I, Karpiskova R, Faldynova M, et al. (1998) Computer-assisted restriction endonuclease analysis of plasmid DNA in field strains of Salmonella Enteritidis. Can J Microbiol 44: 1183-1185. doi: 10.1139/w98-112
|
| [43] |
Rychlik I, Svestkova A, Karpiskova R (2000) Subdivision of Salmonella enterica serovar enteritidis phage types PT14b and PT21 by plasmid profiling. Vet Microbiol 74: 217-225. doi: 10.1016/S0378-1135(00)00185-1
|
| [44] | Cieslik A, Brown D, Paciorek J, et al. (2001) Phage types, plasmid profiles and chromosomal restriction profiles of Salmonella enterica subsp. enterica ser. Enteritidis (S. Enteritidis) isolated in Poland in 1999–2000. Med Dosw Microbiol 53: 185-196. |
| [45] |
Desai M, Threlfall EJ, Stanley J (2001) Fluorescent amplified-fragment length polymorphism subtyping of the Salmonella enterica serovar Enteritidis phage type 4 clone complex. J Clin Microbiol 39: 201-206. doi: 10.1128/JCM.39.1.201-206.2001
|
| [46] |
Nauerby B, Pedersen K, Dietz HH, et al. (2000) Comparison of Danish isolates of Salmonella enterica serovar enteritidis PT9a and PT11 from hedgehogs (Erinaceus europaeus) and humans by plasmid profiling and pulsed-field gel electrophoresis. J Clin Microbiol 38: 3631-3635. doi: 10.1128/JCM.38.10.3631-3635.2000
|
| [47] |
Ling JM, Koo IC, Kam KM, et al. (1998) Antimicrobial susceptibilities and molecular epidemiology of Salmonella enterica serotype Enteritidis strains isolated in Hong Kong from 1986 to 1996. J Clin Microbiol 36: 1693-1699. doi: 10.1128/JCM.36.6.1693-1699.1998
|
| [48] |
Su LH, Chiu CH, Wu TL, et al. (2002) Molecular epidemiology of Salmonella enterica serovar Enteritidis isolated in Taiwan. Microbiol Immunol 46: 833-840. doi: 10.1111/j.1348-0421.2002.tb02770.x
|
| [49] | Shaginyan IA (2000) Role and significance of molecular methods in epidemiological analysis of nosocomial infections. Clin Microbiol Antimicrob Chemother 3: 82-95. |
| [50] | Khazenson LB, Poplavskaia ZhV, Kariagina EI, et al. (1996) Epidemiological data on salmonellosis due to Salmonella enteritidis in some areas of the Russian Federation. J Microbiol Epidemiol Immunobiol 4: 53-57. |
| [51] |
Rodrigue DC, Tauxe RV, Rowe B (1990) International increase in Salmonella Enteritidis: a new pandemic? Epidemiol Infect 105: 21-27. doi: 10.1017/S0950268800047609
|
| [52] |
Bäumler AJ, Hargis BM, Tsolis RM (2000) Tracing the origins of Salmonella outbreaks. Science 287: 50-52. doi: 10.1126/science.287.5450.50
|
| [53] | Rakov AV, Shubin FN, Ivanis VA, et al. (2001) Comparative characterization of infections caused by Salmonella enteritidis of different plasmid profiles. Epidemiol Infect Dis 5: 50-54. |
| [54] | Shubin FN, Rakov AV, Kuznetsova NA, et al. (2006) Structure of Salmonella enteritidis population in Primorye Region on the plasmid analysis data. J Microbiol Epidemiol Immunobiol 3: 28-32. |
| [55] | Shubin FN, Rakov AV, Kuznetsova NA (2011) Microbiological molecular genetic monitoring of enteric infection pathogens as an element of epidemiological surveillance. Bulletin SB RAMS 31: 99-105. |
| [56] | Rakov AV, Shubin FN, Kuznetsova NA (2013) Heterogeneity of 1.4 MDa plasmids in Salmonella enteritidis strains. Bulletin SB RAMS 33: 10-15. |
| [57] |
Shubin FN, Rakov AV, Kuznetsova NA, et al. (2017) Formation of population morbidity with salmonellosis caused by Salmonella enteritidis in regions with incomplete supply of local poultry products. J Microbiol Epidemiol Immunobiol 1: 61-67. doi: 10.36233/0372-9311-2017-1-61-67
|
| [58] | Shubin FN, Kuznetsova NA, Rakov AV, et al. (2018) Specific features of molecular epidemiology of imported morbidity caused by Salmonella Enteritidis introduced strains with specific plasmid types. Epidemiol Infect Dis 23: 73-76. |
| [59] |
Rakov AV, Kuznetsova NA, Solovyeva AS, et al. (2018) Cluster analysis of Salmonella Enteritidis isolated in the Siberia and Far East of Russia. Pacific Med J 4: 23-26. doi: 10.17238/PmJ1609-1175.2018.4.23-26
|
| [60] | Rakov AV, Shubin FN, Kuznetsova NA, et al. (2019) Heterogeneity of 2.3 MDa plasmids in Salmonella Enteritidis strains. Siberian Sci Med J 39: 40-45. |
| [61] | Kuznetsova NA, Solovyeva AS, Rakov AV (2018) Antibiotic resistance of Salmonella Enteritidis strains, circulated in territory of the Siberia and Far East, at multi-year monitoring. Health Med Ecol Sci 3: 50-58. |
| [62] |
Rakov AV, Shubin FN, Kuznetsova NA, et al. (2016) Estimated microbiological and epidemiological of the outbreak of salmonellosis in the child care institution in Murmansk. Health Med Ecol Sci 4: 26-30. doi: 10.4103/2321-4848.183351
|
| [63] |
Döpfer D, Buist W, Soyer Y, et al. (2008) Assessing genetic heterogeneity within bacterial species isolated from gastrointestinal and environmental samples: how many isolates does it take? Appl Environ Microbiol 74: 3490-3496. doi: 10.1128/AEM.02789-07
|
| [64] | Timchenko NF, Rakov AV, Terentyeva NA, et al. (2019) Characteristics of the mixed bacteria of the Enterobacteriaceae family Yersinia pseudotuberculosis and Salmonella Enteritidis in vitro. Health Med Ecol Sci 1: 19-22. |
| [65] | Yakovlev AA, Pozdeeva ES (2018) Possible mechanisms of self-regulation of parasitic systems in the biogeocenosis. Vestnik RAMS 73: 195-205. |

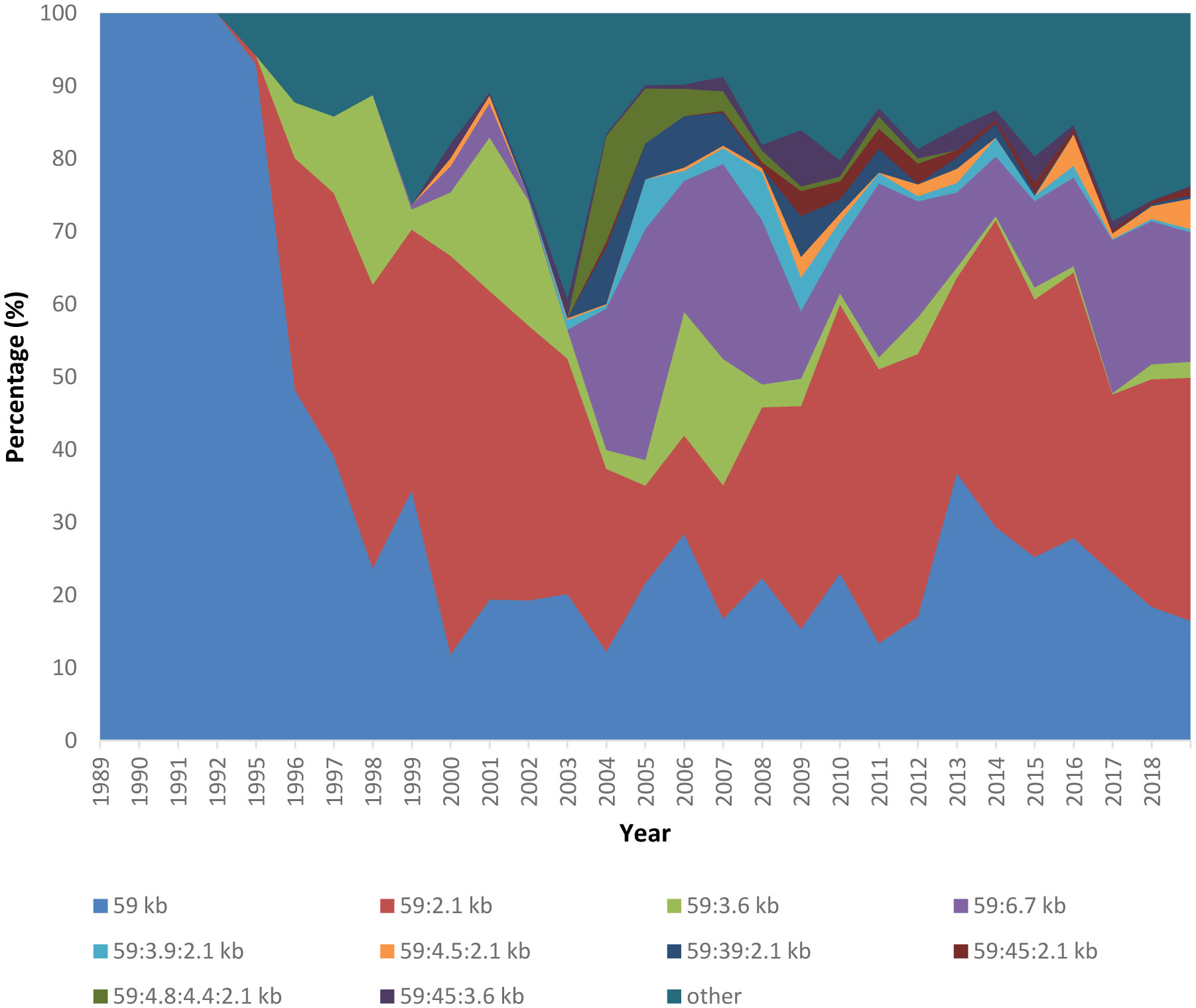
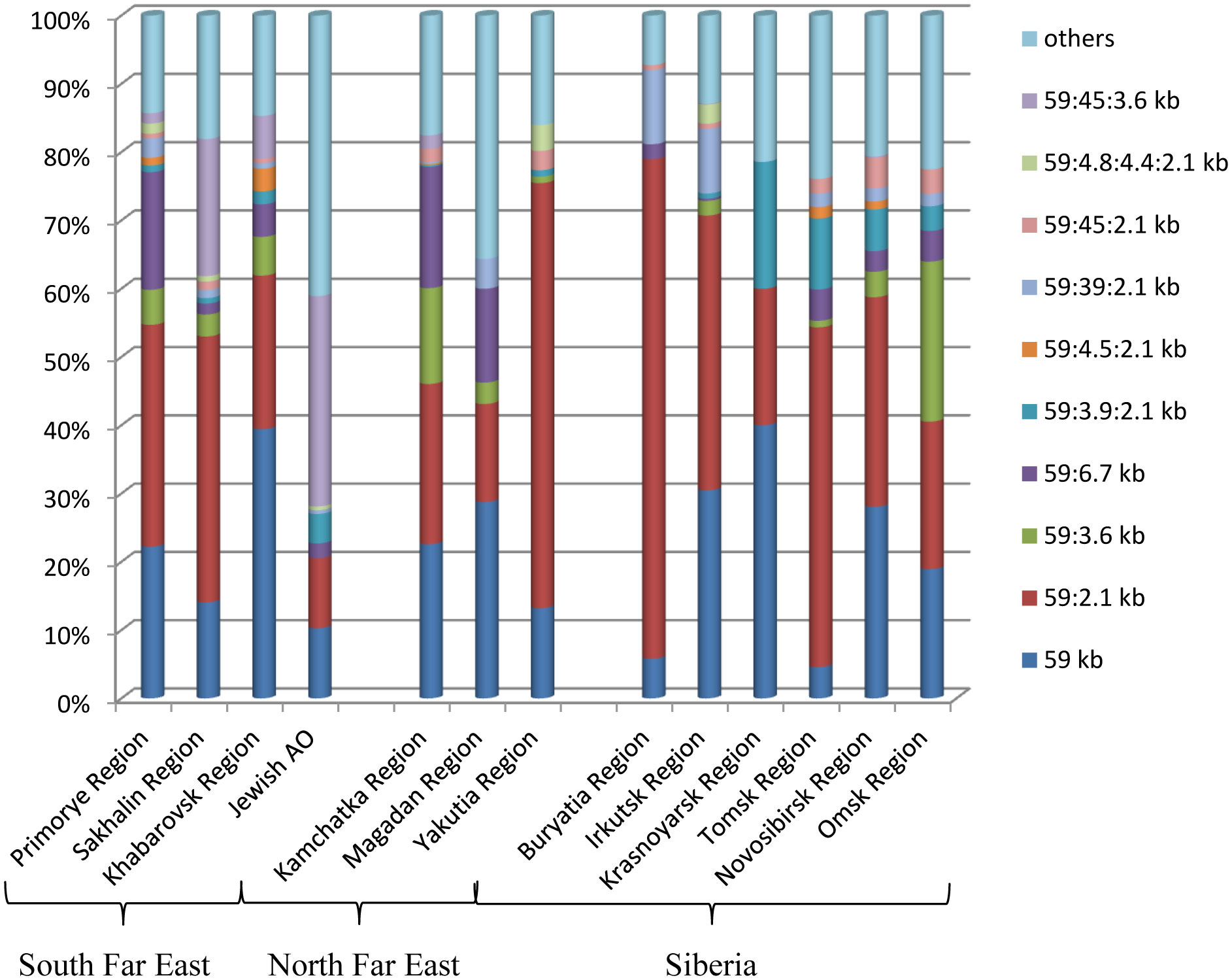
Figures(3)

Alexey V. Rakov, Natalya A. Kuznetsova, Anatoly A. Yakovlev. Genetic diversity of Salmonella enterica subsp. enterica serovar Enteritidis in the Siberia and Far East of Russia based on plasmid profiles[J]. AIMS Microbiology, 2020, 6(2): 106-120. doi: 10.3934/microbiol.2020007







 DownLoad:
DownLoad: