Citation: Temitayo O. Olowomofe, J. O. Oluyege, B.I. Aderiye, O. A. Oluwole. Degradation of poly aromatic fractions of crude oil and detection of catabolic genes in hydrocarbon-degrading bacteria isolated from Agbabu bitumen sediments in Ondo State[J]. AIMS Microbiology, 2019, 5(4): 308-323. doi: 10.3934/microbiol.2019.4.308

| [1] | Oboh BO, Ilori OM, Akinyemi JO, et al. (2006) Hydrocarbon degrading potentials of bacteria isolated from a Nigeria Bitumen (Tarsand) deposit. Nat Sci 4: 51–57. |
| [2] | Kadafa AA (2012) Environmental impacts of oil exploration and exploitation in the Niger Delta of Nigeria. Global J Sci Front Res Environ Earth Sci 12: 19–28. |
| [3] | F M E. (2006) Niger Delta natural resource damage assessment and restoration project: Phase 1-Scoping report. Res J Environ Toxicol 10: 29–37. |
| [4] | Bidoia ED, Montagnolli RN, Lopes PRM (2010) Microbial biodegradation potential of hydrocarbons evaluated by colorimetric technique: a case study, In: Mendez-Vilas A, Current research, technology and education topics in applied microbiology and microbial biotechnology, FORMATEX, Badajoz, Spain, 40–45. |
| [5] |
Mwaura AN, Mbatia BN, Muge EK, et al. (2018) Screening and characterization of hydrocarbonoclastic bacteria isolated from oil-contaminated soils from auto garages. Int J Microbiol Biotechn 3: 11–24. doi: 10.11648/j.ijmb.20180301.13

|
| [6] |
Boonchan S, Britz ML, Stanley GA (2000) Degradation and mineralization of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial co-cultures. Appl Environ Microbiol 66: 1007–1019. doi: 10.1128/AEM.66.3.1007-1019.2000
|
| [7] |
Mao J, Luo Y, Teng Y, et al. (2012) Bioremediation of polycyclic aromatic hydrocarbon-contaminated soil by a bacterial consortium and associated microbial community changes. Int Biodeterior Biodegrad 70: 141–147. doi: 10.1016/j.ibiod.2012.03.002
|
| [8] | Lam SS, Russell AD, Lee CL, et al. (2012) Production of hydrogen and light hydrocarbons as a potential gaseous fuel from microwave-heated pyrolysis of waste automotive engine oil. Int J Hydrogen Energ 30: 5011–5021. |
| [9] |
Islas-garcia A, Vega-loyo L, Aguilar-lopez R, et al. (2015) Evaluation of hydrocarbons and organochlorine pesticides and their tolerant microorganisms from an agricultural soil to define its bioremediation feasibility. J Environ Sci Health B 50: 99–108. doi: 10.1080/03601234.2015.975605
|
| [10] |
Watanabe K (2001) Microorganisms relevant to bioremediation. Curr Opin Biotechnol 12: 231–241. doi: 10.1016/S0958-1669(00)00204-4
|
| [11] | Bushnell LD, Haas HF (1941) The utilization of hydrocarbons by microorganisms. J Bacteriol 41: 653–673. |
| [12] | Oluwole OA, Oluyege JO, Odeyemi AT, et al. (2017) Growth patterns and degradative potentials of Pseudomonas sp. isolated from waste dumpsite soil in crude oil supplemented soil extract and mineral salts media. J Adv Biolo Biotechnol 12: 1–23. |
| [13] |
Margesin R, Schinner F (2001) Bioremediation (natural attenuation and biostimulation) of diesel-oil-contaminated soil in Alpine glacier sking area. Appl Environ Microbiol 67: 3127–3133. doi: 10.1128/AEM.67.7.3127-3133.2001
|
| [14] |
Semple KT, Morriss A, Paton GI (2003) Bioavailability of hydrophobic organic contaminants in soils: fundamental concepts and techniques for analysis. Eur J Soil Sci 54: 809–818. doi: 10.1046/j.1351-0754.2003.0564.x
|
| [15] | Sathishkumar M, Arthur RB, Sang-Ho B, et al. (2008) Biodegradation of crude oil by individual bacterial strains and a mixed bacterial consortium isolated from hydrocarbon contaminated areas. Clean 36: 92–96. |
| [16] | Obayori OS, Ilori MO, Adebusoye SA, et al. (2009b) Degradation of hydrocarbons and biosurfactant production by Pseudomonas sp. strain LP1. World J Microb Biot 25: 1615–1623. |
| [17] |
Olabemiwo MO, Adediran GO, Adekola FA, et al. (2014) Biodegradation of hydrocarbon compounds in Agbabu natural bitumen. Afr J Biotechnol 13: 1257–1264. doi: 10.5897/AJB2013.12974
|
| [18] | Olowomofe Temitayo O, Oluyege JO, Olawole OA, et al. (2018) Catechol-2,3-dioxygenase and Lipase Activities during Degradation of Crude Oil by Hydrocarbon-degrading Bacteria Isolated from Bitumen-polluted Surface Water in Agbabu, Ondo State. Int J Environ Biorem Biodegrad 6: 18–25. |
| [19] | Olowomofe TO, Oluyege JO, Sowole DO (2017) Isolation, screening and characterization of hydrocarbon-utilizing bacteria isolated from bitumen-contaminated surface water in Agbabu, Ondo State. J Adv Biol Biotechnol 15: 1–9. |
| [20] | Orhorhoro E, Effiong E, Abu G (2018) Laboratory-scale bioremediation of crude oil polluted soil using a consortia of rhizobacteria obtained from plants in Gokana-Ogoni, Rivers State. J Adv Microbiol 9: 1–17. |
| [21] | Minai-Tehrani D, Herfatmanesh A (2007) Biodegradation of aliphatic and aromatic fractions of heavy crude oil-contaminated soil: a pilot study. J Biodegrad Biorem 11: 71–76. |
| [22] |
Baek KH, Yoon BD, Oh HM, et al. (2006) Biodegradation of aliphatic and aromatic hydrocarbons by Nocardia sp. H17–1. Geomicrobiology J 23: 253–259. doi: 10.1080/01490450600760633
|
| [23] | Tancsics A, Szabó I, Baka E, et al. (2008) Investigation of catechol 2,3-dioxygenase and 16S rRNA gene diversity in hypoxic, petroleum hydrocarbon contaminated groundwater. Syst Appl Microbiol 33: 398–406. |
| [24] | Khan MA, Biswas B, Smith E, et al. (2018) Toxicity assessment of fresh and weathered petroleum hydrocarbons in contaminated soil. Chemosphere 8: 35–42. |
| [25] | Aurepatipan N, Champreda V, Kanokratana P, et al. (2018) Assessment of bacterial communities and activities of thermotolerant enzymes produced by bacteria indigenous to oil- bearing sandstone cores for potential application in Enhanced Oil Recovery. J Petroleum Sci Eng163: 295–302. |
| [26] |
Ji H, Gong Y, Duan J, et al. (2018) Degradation of petroleum hydrocarbons in seawater by simulated surface level atmospheric ozone: reaction kinetics and effect of oil dispersant. Mar Pollut Bull 135: 427–440. doi: 10.1016/j.marpolbul.2018.07.047
|
| [27] |
Gupta G, Kumar V, Pal AK (2016) Biodegradation of polycyclic aromatic hydrocarbons by microbial consortium: a distinctive approach for decontamination of soil. Soil Sediment Contam 25: 597–623. doi: 10.1080/15320383.2016.1190311
|
| [28] |
Das K, Mukherjee F (2007) Crude petroleum-oil biodegradation efficiency of Bacillus subtilis and Pseudomonas aeruginosa strains isolated from a petroleum-oil contaminated soil from North-East India. Bioresour Technol 98: 1339–1345. doi: 10.1016/j.biortech.2006.05.032
|
| [29] |
Amini S, Zulkify AH, et al. (2011) Molecular identification and characterization of a bacterium that has potential to degrade low concentration of halo alkanoic acid. Res J Microbiol 6: 552–559. doi: 10.3923/jm.2011.552.559
|
| [30] |
Olabemiwo OM, Adediran GO, Adekola FA, et al. (2011) Impact of simulated Agbabu bitumen Leachate on haematological and biochemical parameters of wistar albino rat. Res J Environ Toxicol 5: 213–221. doi: 10.3923/rjet.2011.213.221
|
| [31] | International Agency for Research on Cancer (IARC) (2010) Some non-heterocyclic polycyclic aromatic hydrocarbons and some related exposures. Int Agency Res Cancer 92: 45–66. |
| [32] | Deng D, Li C, Ju Q, et al. (1999) Systematic extensive laboratory studies of microbial EOR mechanisms and microbial EOR application results in Changing Oilfield. SPE Asia Pacific Oil and Gas Conference and Exhibition; Society of Petroleum Engineers, 56. |
| [33] |
Etoumi A, Musrati IE, Gammoudi BE, et al. (2008) The reduction of wax precipitation in waxy crude oils by Pseudomonas species. J Ind Microbiol Biotechnol 35: 1241–1245. doi: 10.1007/s10295-008-0420-z
|
| [34] |
Al-Sayegh A, Al-Wahaibi Y, Al-Bahry S, et al. (2015) Microbial enhanced heavy crude oil recovery through biodegradation using bacterial isolates from an Omani oil field. Microb Cell Fact 14: 141. doi: 10.1186/s12934-015-0330-5
|
| [35] |
Kumari S, Regar RK, Manickam N (2018) Improved polycyclic aromatic hydrocarbon degradation in a crude oil by individual and a consortium of bacteria. Bioresour Technol 254: 174–179. doi: 10.1016/j.biortech.2018.01.075
|
| [36] |
Anastasi A, Coppola T, Prigione V (2009) Pyrene degradation and detoxification in soil by a consortium of basidiomycetes isolated from compost: Role of laccases and peroxidases. J Hazard Mater 165: 1229–1233. doi: 10.1016/j.jhazmat.2008.10.032
|
| [37] |
Wu M, Chen L, Tian Y, et al. (2013) Degradation of polycyclic aromatic hydrocarbons by microbial consortia enriched from three soils using two different culture media. Environ Pollut 178: 152–158. doi: 10.1016/j.envpol.2013.03.004
|
| [38] |
Zafra G, Absalón AE, Cortés-Espinosa DV (2015) Morphological changes and growth of filamentous fungi in the presence of high concentrations of PAHs. Braz J Microbiol 46: 937–941. doi: 10.1590/S1517-838246320140575
|
| [39] |
Wanapaisan P, Laothamteep N, Vejarano F, et al. (2018). Synergistic degradation of pyrene by five culturable bacteria in a mangrove sediment-derived bacterial consortium. J Hazard Mater 342: 561–570. doi: 10.1016/j.jhazmat.2017.08.062
|
| [40] |
Ghorbannezhad H, Moghimi H, Taheri RA (2018) Enhanced biodegradation of phenol by magnetically immobilized Trichosporon cutaneum. Ann Microbiol 68: 485–491. doi: 10.1007/s13213-018-1353-z
|
| [41] |
Owsianiak M., Szulc A, Chrzanowski L, et al. (2009) Biodegradation and surfactant-mediated biodegradation of diesel fuel by 218 microbial consortia are not correlated to cell surface hydrophobicity. Appl Microbiol Biot 84: 545–553. doi: 10.1007/s00253-009-2040-6
|
| [42] | Vinithini C, Sudhakar S, Ravikumar R (2015) Biodegradation of petroleum and crude oil by Pseudomonas putida and Bacillus cereus. Int J Curr Microbiol Appl Sci 4: 318–329. |
| [43] | Ma Y, Wang L, Shao Z (2006) Pseudomonas, the dominant polycyclic aromatic hydrocarbon- degrading bacteria isolated from Antarctic soils and the role of large plasmids in horizontal gene transfer. Natl Cent Biotechnol Inf 8: 455–65. |
| [44] | Jyothi KK, Surendra B, Nancy CK, et al. (2012) Identification and Isolation of Hydrocarbon- degrading Bacteria by Molecular Characterization. Helix 2: 105–111. |
| [45] |
El-Sayed WS (2006) Molecular cloning of gene nahH encoding extradiol-type dioxygenase from the NAH plasmid of Pseudomonas stutzeri NA1. Ann Microbiol 56: 89. doi: 10.1007/BF03174987
|
| [46] | Veenagayathri K, Vasudevan N (2011) Ortho and meta cleavage dioxygenases detected during the degradation of phenolic compounds by a moderately halophilic bacterial consortium. Int Res J Microbiol 2: 406–414. |
| [47] | Zhao K, Guo X, Gong JA (2013) Novel benzoate-degrading Rhodococcus strain contains three catA genes with one being transcriptionally active during the growth on benzoate. Res J Environ Earth Sci 34: 401–407. |
| [48] | Dokic L, Narancic T, Nikodinovic-Runic J, et al. (2011) Four Bacillus sp. soil isolates capable of degrading phenol, toluene, biphenyl, naphthalene and other aromatic compounds exhibit different aromatic catabolic potentials. Arch Biolo Sci 63: 1057–1067. |
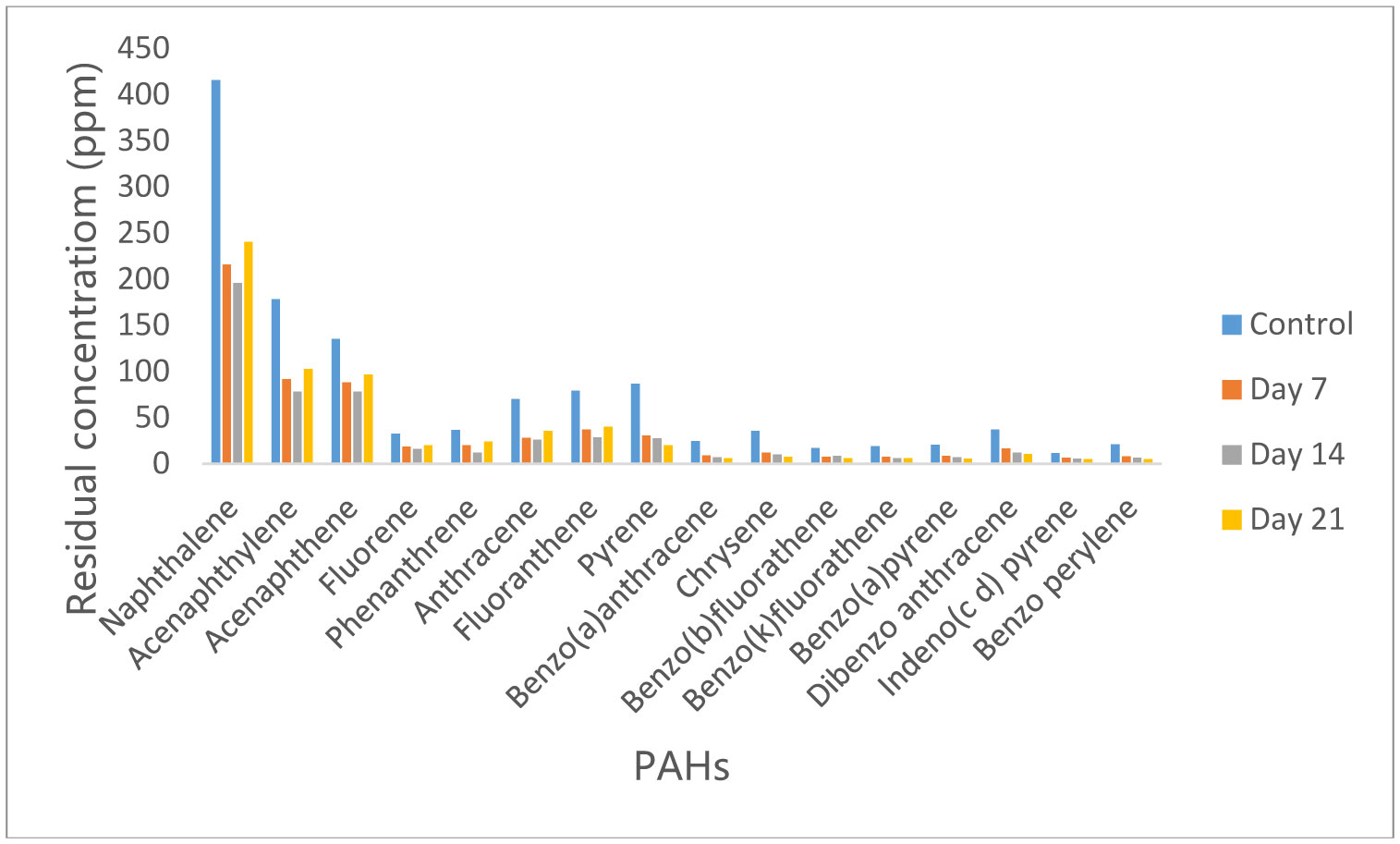
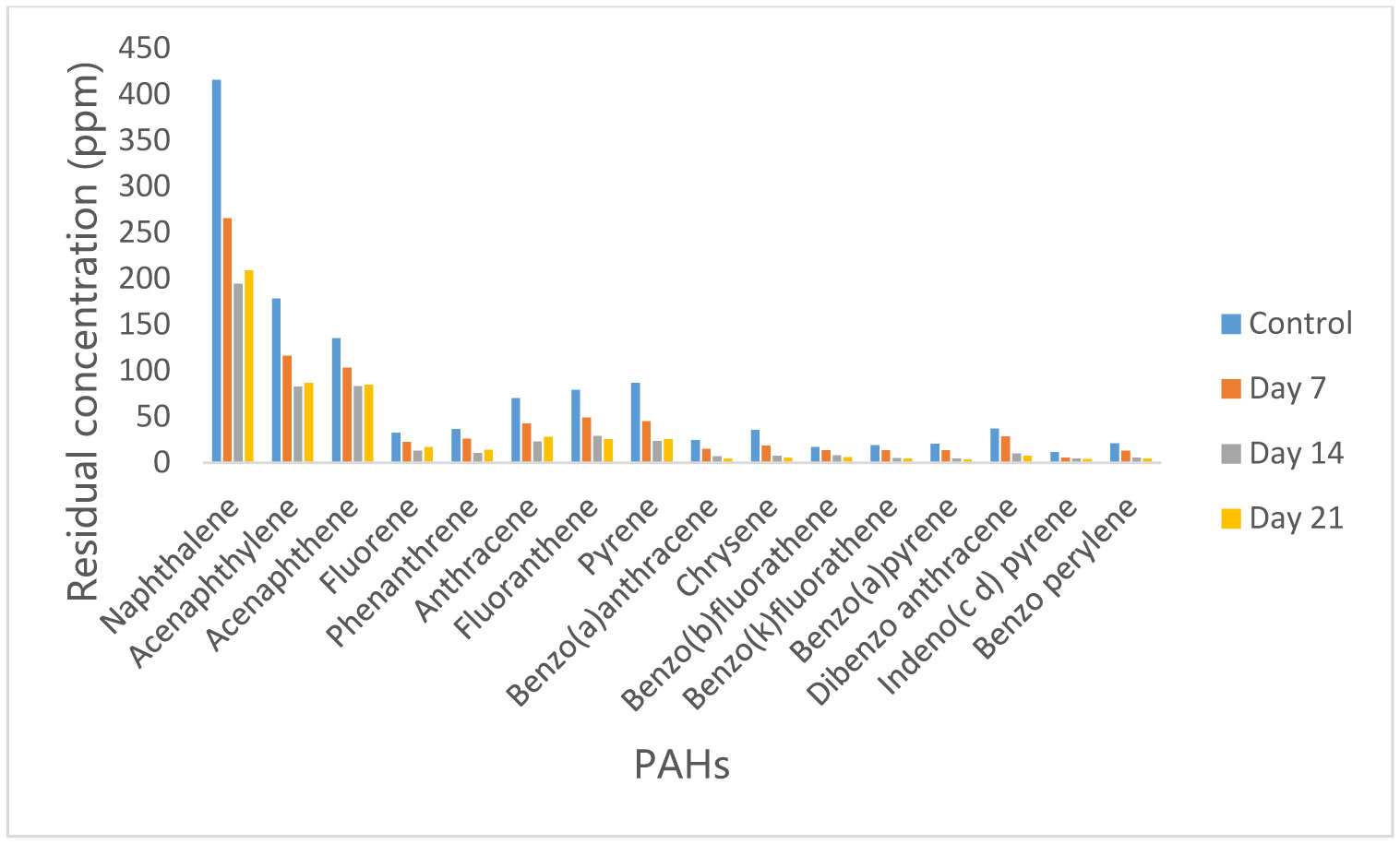
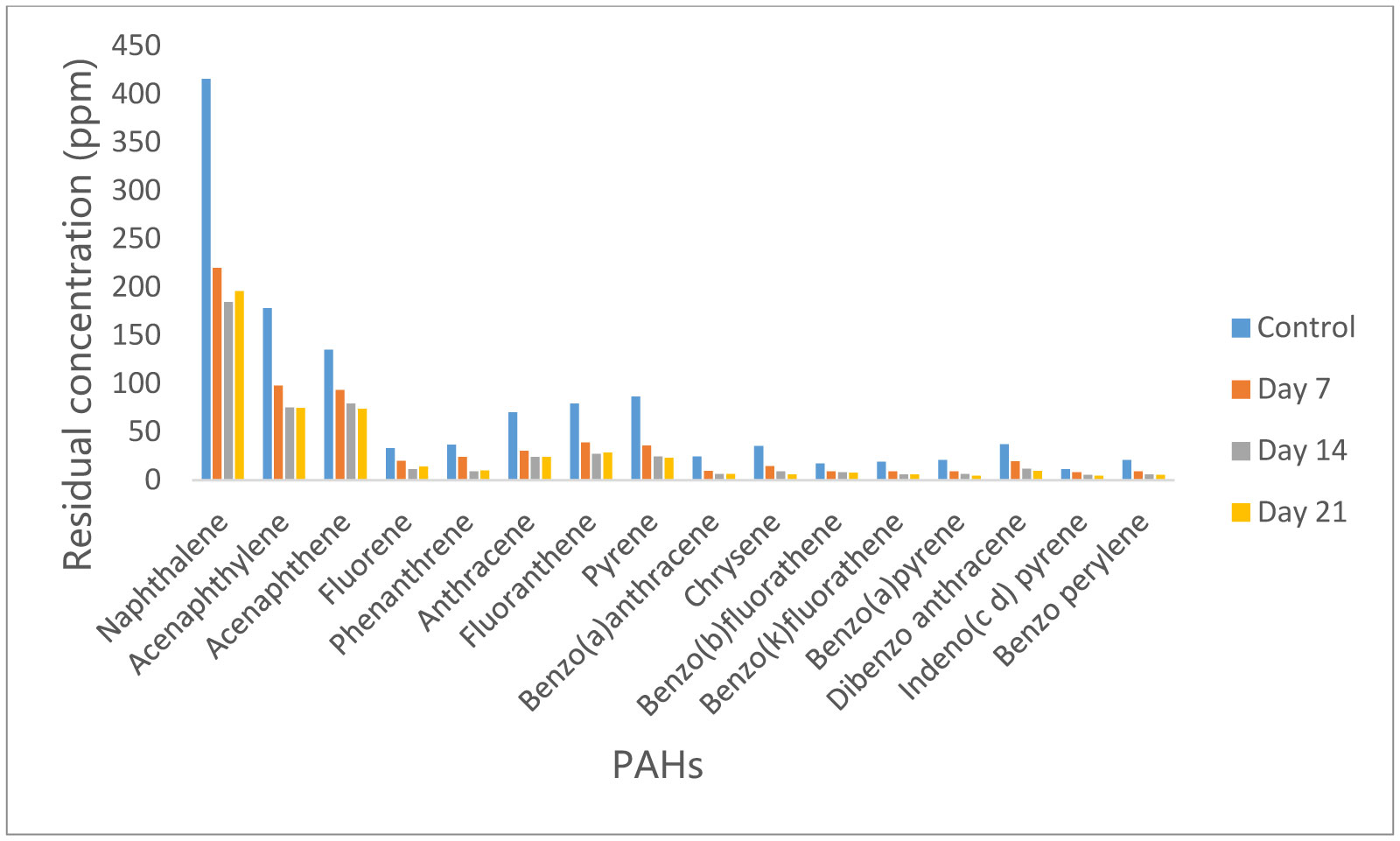
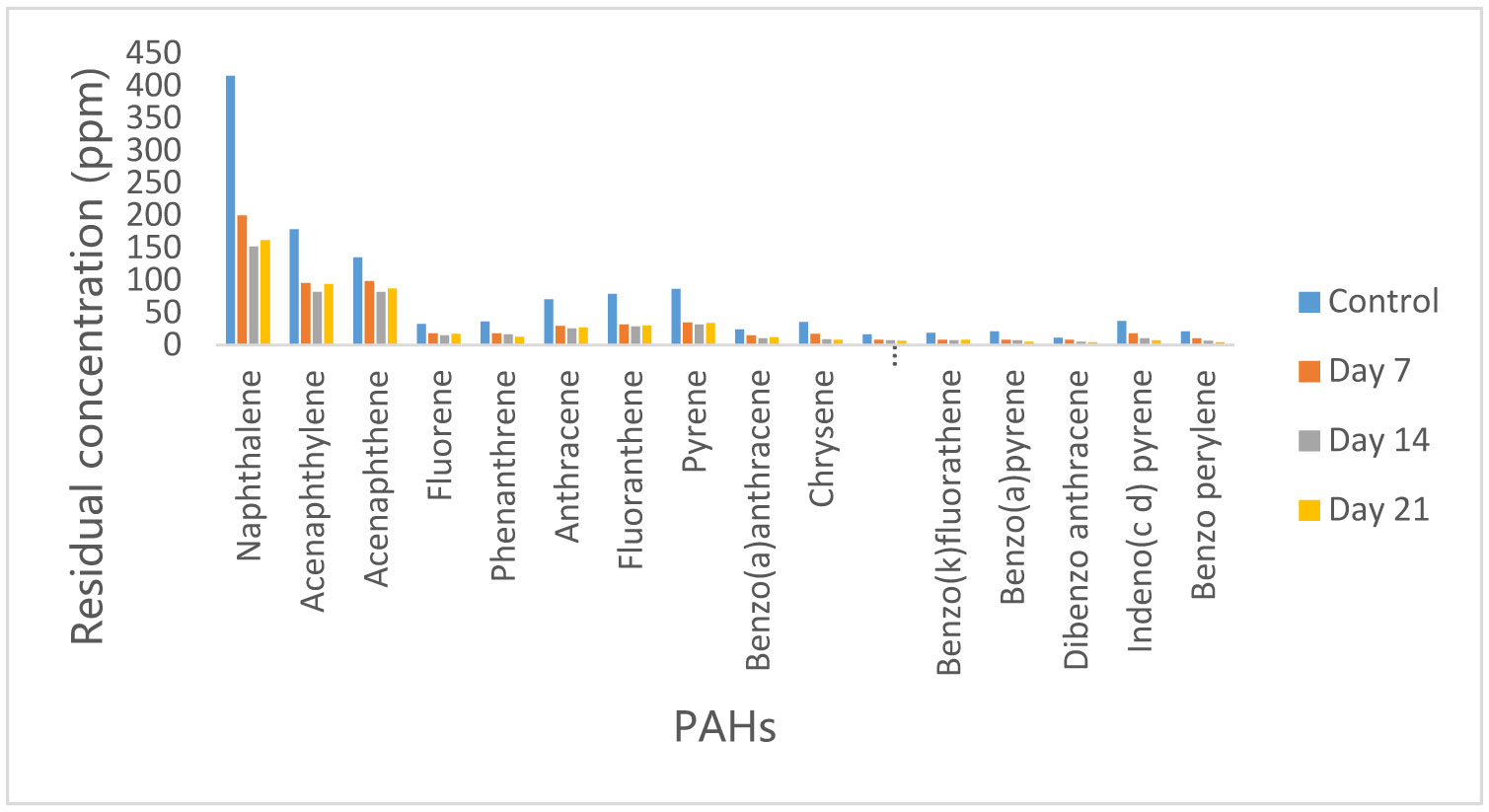
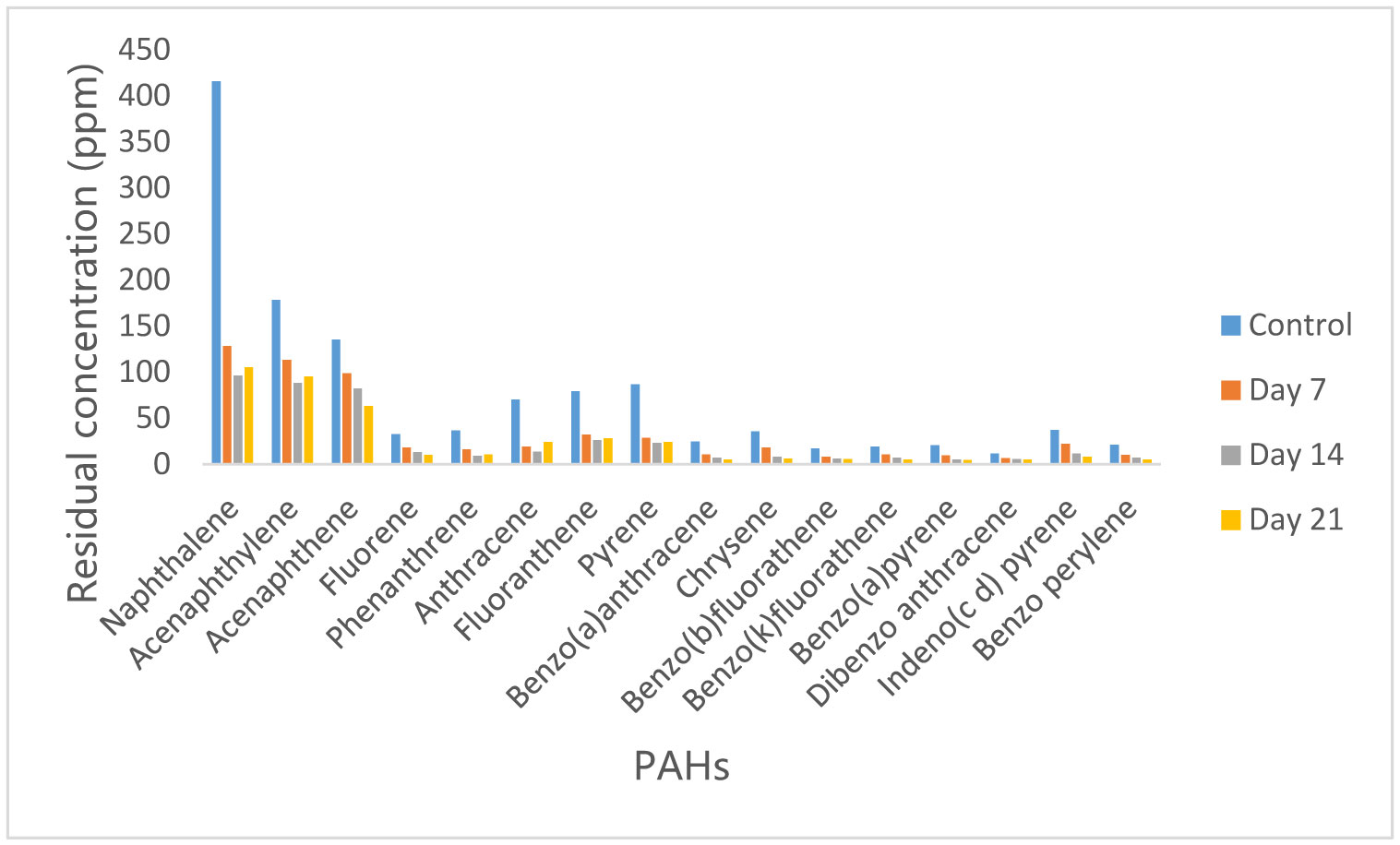
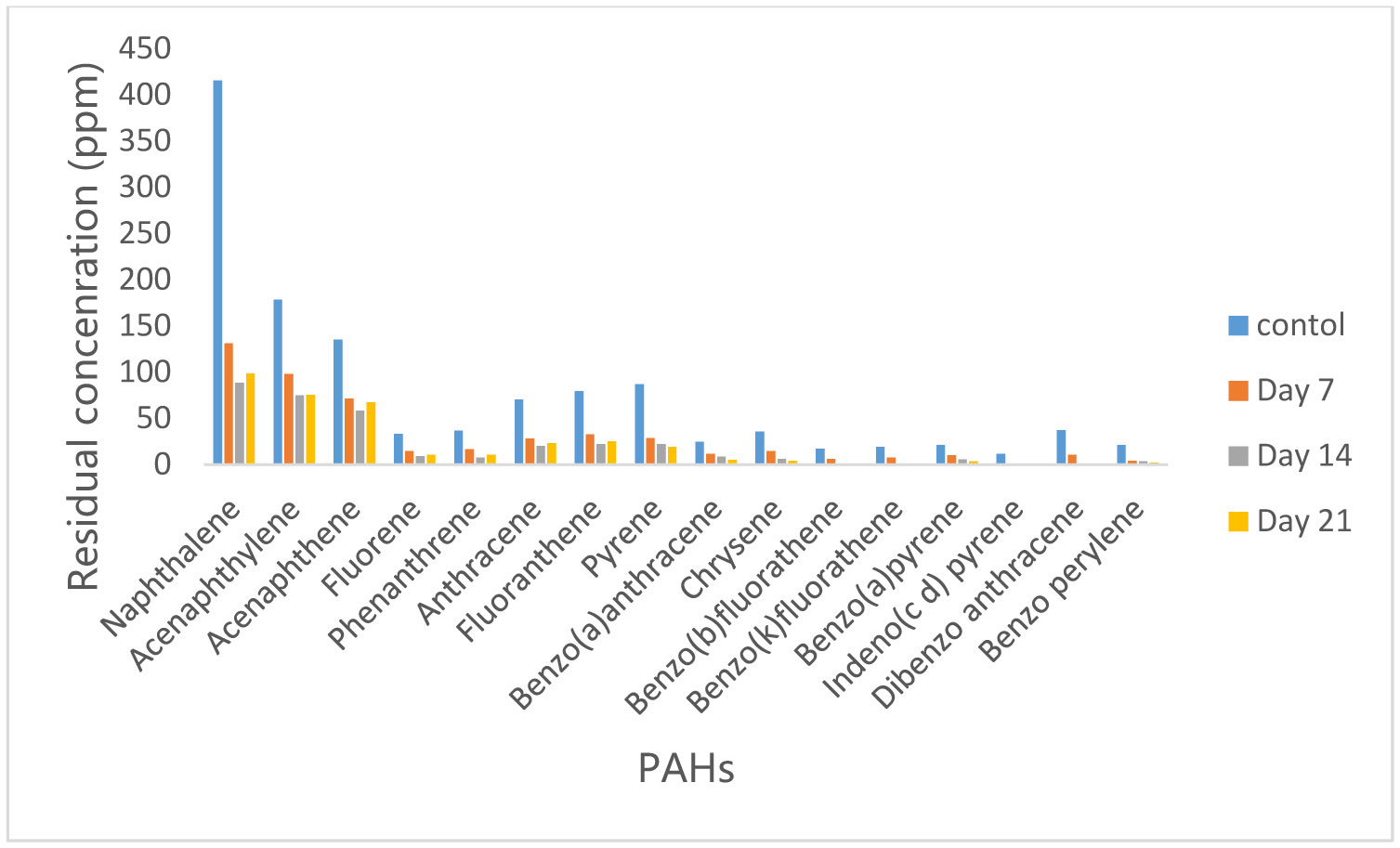
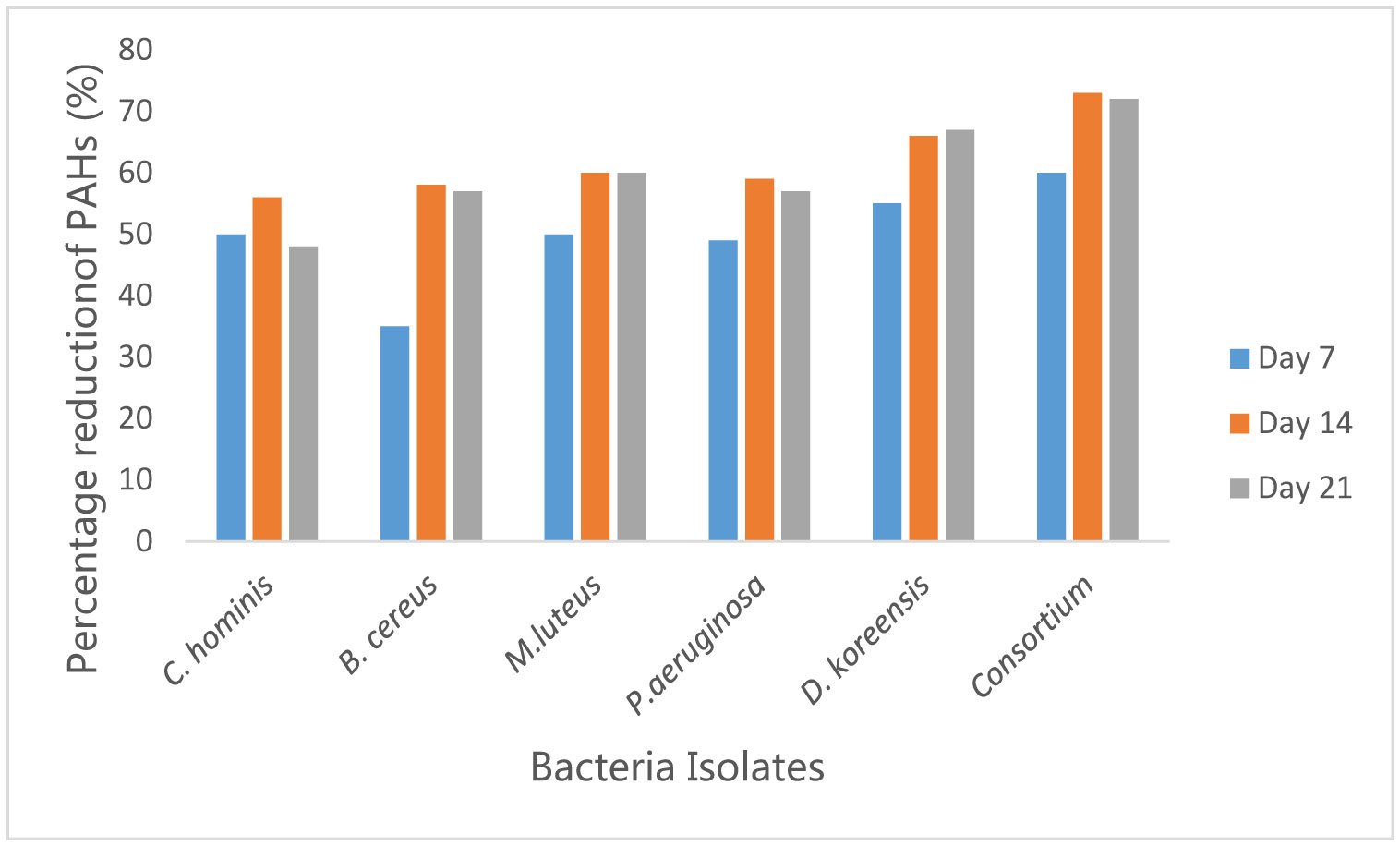
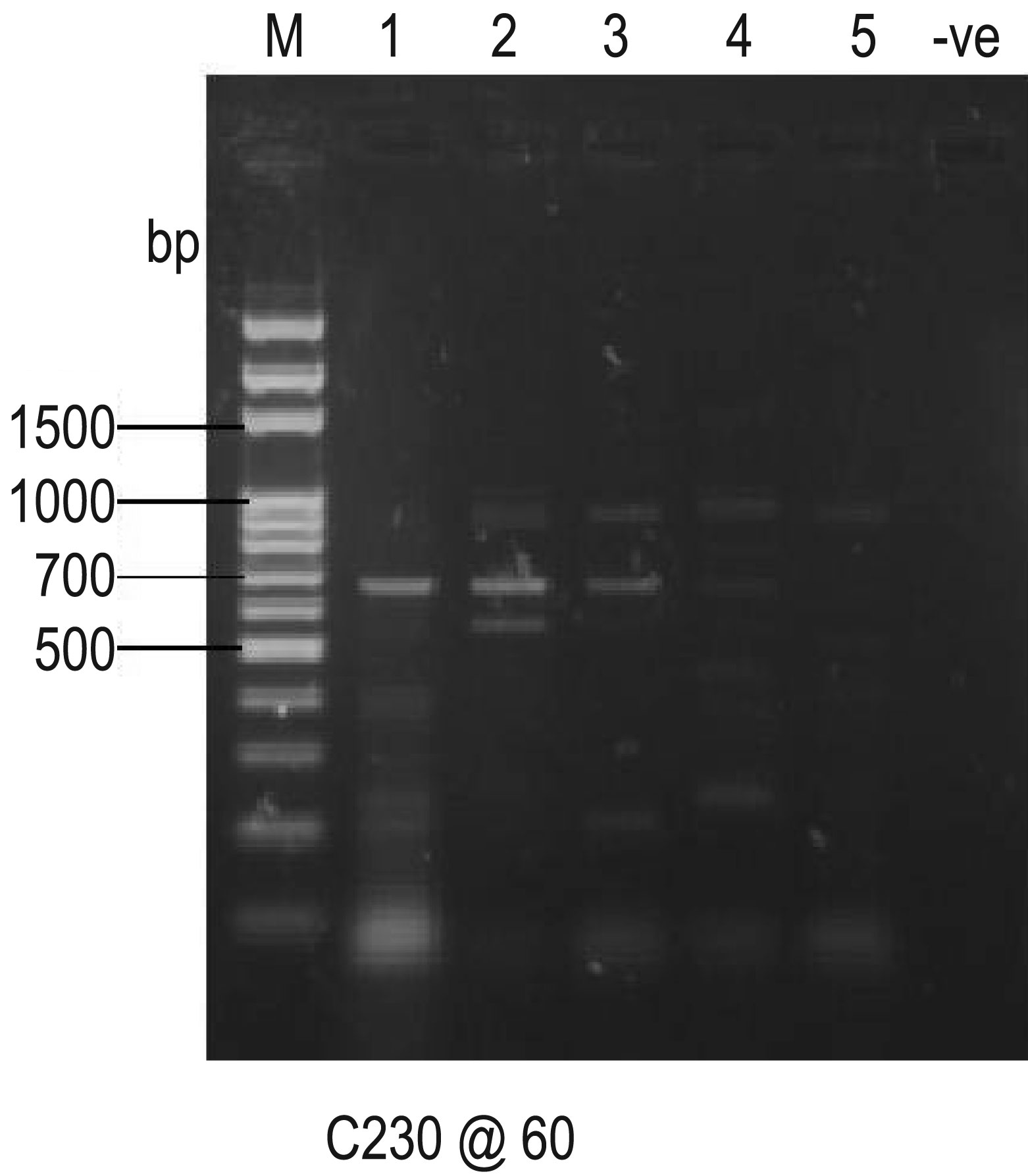
Figures(10)

Temitayo O. Olowomofe, J. O. Oluyege, B.I. Aderiye, O. A. Oluwole. Degradation of poly aromatic fractions of crude oil and detection of catabolic genes in hydrocarbon-degrading bacteria isolated from Agbabu bitumen sediments in Ondo State[J]. AIMS Microbiology, 2019, 5(4): 308-323. doi: 10.3934/microbiol.2019.4.308







 DownLoad:
DownLoad: