Citation: Angela Tramonti, Fiorenzo De Santis, Eugenia Pennacchietti, Daniela De Biase. The yhiM gene codes for an inner membrane protein involved in GABA export in Escherichia coli[J]. AIMS Microbiology, 2017, 3(1): 71-87. doi: 10.3934/microbiol.2017.1.71

| [1] |
Lund P, Tramonti A, De Biase D (2014) Coping with low pH: molecular strategies in neutralophilic bacteria. FEMS Microbiol Rev 38: 1091–1125. doi: 10.1111/1574-6976.12076

|
| [2] |
Foster JW (2004) Escherichia coli acid resistance: tales of an amateur acidophile. Nat Rev Microbiol 2: 898–907. doi: 10.1038/nrmicro1021
|
| [3] |
De Biase D, Lund PA (2015) The Escherichia coli Acid Stress Response and Its Significance for Pathogenesis. Adv Appl Microbiol 92: 49–88. doi: 10.1016/bs.aambs.2015.03.002
|
| [4] | Lin J, Smith MP, Chapin KC, et al. (1996) Mechanisms of acid resistance in enterohemorrhagic Escherichia coli. Appl Environ Microbiol 62: 3094–3100. |
| [5] |
De Biase D, Tramonti A, Bossa F, et al. (1999) The response to stationary-phase stress conditions in Escherichia coli: role and regulation of the glutamic acid decarboxylase system. Mol Microbiol 32: 1198–1211. doi: 10.1046/j.1365-2958.1999.01430.x
|
| [6] | Castanie-Cornet MP, Penfound TA, Smith D, et al. (1999) Control of acid resistance in Escherichia coli. J Bacteriol 181: 3525–3535. |
| [7] |
Eguchi Y, Ishii E, Hata K, et al. (2011) Regulation of acid resistance by connectors of two-component signal transduction systems in Escherichia coli. J Bacteriol 193: 1222–1228. doi: 10.1128/JB.01124-10
|
| [8] | Burton NA, Johnson MD, Antczak P, et al. (2010) Novel aspects of the acid response network of E. coli K-12 are revealed by a study of transcriptional dynamics. J Mol Biol 401: 726–742. |
| [9] | Tramonti A, De Canio M, De Biase D (2008) GadX/GadW-dependent regulation of the Escherichia coli acid fitness island: transcriptional control at the gadY-gadW divergent promoters and identification of four novel 42 bp GadX/GadW-specific binding sites. Mol Microbiol 70: 965–982. |
| [10] |
Sayed AK, Foster JW (2009) A 750 bp sensory integration region directs global control of the Escherichia coli GadE acid resistance regulator. Mol Microbiol 71: 1435–1450. doi: 10.1111/j.1365-2958.2009.06614.x
|
| [11] |
Mates AK, Sayed AK, Foster JW (2007) Products of the Escherichia coli acid fitness island attenuate metabolite stress at extremely low pH and mediate a cell density-dependent acid resistance. J Bacteriol 189: 2759–2768. doi: 10.1128/JB.01490-06
|
| [12] |
De Biase D, Pennacchietti E (2012) Glutamate decarboxylase-dependent acid resistance in orally acquired bacteria: function, distribution and biomedical implications of the gadBC operon. Mol Microbiol 86: 770–768. doi: 10.1111/mmi.12020
|
| [13] |
Tsai MF, McCarthy P, Miller C (2013) Substrate selectivity in glutamate-dependent acid resistance in enteric bacteria. Proc Natl Acad Sci USA 110: 5898–5902. doi: 10.1073/pnas.1301444110
|
| [14] |
Ma D, Lu P, Shi Y (2013) Substrate selectivity of the acid-activated glutamate/gamma-aminobutyric acid (GABA) antiporter GadC from Escherichia coli. J Biol Chem 288: 15148–15153. doi: 10.1074/jbc.M113.474502
|
| [15] |
Lu P, Ma D, Chen Y, et al. (2013) L-glutamine provides acid resistance for Escherichia coli through enzymatic release of ammonia. Cell Res 23: 635–644. doi: 10.1038/cr.2013.13
|
| [16] |
Bordi C, Theraulaz L, Mejean V, et al. (2003) Anticipating an alkaline stress through the Tor phosphorelay system in Escherichia coli. Mol Microbiol 48: 211–223. doi: 10.1046/j.1365-2958.2003.03428.x
|
| [17] |
Hommais F, Krin E, Laurent-Winter C, et al. (2001) Large-scale monitoring of pleiotropic regulation of gene expression by the prokaryotic nucleoid-associated protein, H-NS. Mol Microbiol 40: 20–36. doi: 10.1046/j.1365-2958.2001.02358.x
|
| [18] |
Weber H, Polen T, Heuveling J, et al. (2005) Genome-wide analysis of the general stress response network in Escherichia coli: sigmaS-dependent genes, promoters, and sigma factor selectivity. J Bacteriol 187: 1591–1603. doi: 10.1128/JB.187.5.1591-1603.2005
|
| [19] | Shepherd M, Sanguinetti G, Cook GM, et al. (2010) Compensations for diminished terminal oxidase activity in Escherichia coli: cytochrome bd-II-mediated respiration and glutamate metabolism. J Biol Chem 285: 18464–18472. |
| [20] |
Hayes ET, Wilks JC, Sanfilippo P, et al. (2006) Oxygen limitation modulates pH regulation of catabolism and hydrogenases, multidrug transporters, and envelope composition in Escherichia coli K-12. BMC Microbiol 6: 89. doi: 10.1186/1471-2180-6-89
|
| [21] |
Tucker DL, Tucker N, Ma Z, et al. (2003) Genes of the GadX-GadW regulon in Escherichia coli. J Bacteriol 185: 3190–3201. doi: 10.1128/JB.185.10.3190-3201.2003
|
| [22] |
Tucker DL, Tucker N, Conway T (2002) Gene expression profiling of the pH response in Escherichia coli. J Bacteriol 184: 6551–6558. doi: 10.1128/JB.184.23.6551-6558.2002
|
| [23] |
Eguchi Y, Oshima T, Mori H, et al. (2003) Transcriptional regulation of drug efflux genes by EvgAS, a two-component system in Escherichia coli. Microbiology 149: 2819–2828. doi: 10.1099/mic.0.26460-0
|
| [24] |
Masuda N, Church GM (2002) Escherichia coli gene expression responsive to levels of the response regulator EvgA. J Bacteriol 184: 6225–6234. doi: 10.1128/JB.184.22.6225-6234.2002
|
| [25] |
Kailasan-Vanaja S, Bergholz TM, Whittam TS (2009) Characterization of the Escherichia coli O157:H7 Sakai GadE regulon. J Bacteriol 191: 1868–1877. doi: 10.1128/JB.01481-08
|
| [26] |
Ito A, May T, Kawata K, et al. (2008) Significance of rpoS during maturation of Escherichia coli biofilms. Biotechnol Bioeng 99: 1462–1471. doi: 10.1002/bit.21695
|
| [27] | Nguyen TM, Sparks-Thissen RL (2012) The inner membrane protein, YhiM, is necessary for Escherichia coli (E. coli) survival in acidic conditions. Arch Microbiol 194: 637–641. |
| [28] |
White-Ziegler CA, Davis TR (2009) Genome-wide identification of H-NS-controlled, temperature-regulated genes in Escherichia coli K-12. J Bacteriol 191: 1106–1110. doi: 10.1128/JB.00599-08
|
| [29] |
White-Ziegler CA, Um S, Perez NM, et al. (2008) Low temperature (23 degrees C) increases expression of biofilm-, cold-shock- and RpoS-dependent genes in Escherichia coli K-12. Microbiology 154: 148–166. doi: 10.1099/mic.0.2007/012021-0
|
| [30] |
Yoshida M, Kashiwagi K, Shigemasa A, et al. (2004) A unifying model for the role of polyamines in bacterial cell growth, the polyamine modulon. J Biol Chem 279: 46008–46013. doi: 10.1074/jbc.M404393200
|
| [31] | Ausubel FM, Brent R, Kingston RE, et al. (1987) Current protocols in molecular biology, New York, NY: John Wiley and Sons. |
| [32] | Vogel HJ, Bonner DM (1956) Acetylornithinase of Escherichia coli: partial purification and some properties. J Biol Chem 218: 97–106. |
| [33] |
Datsenko KA, Wanner BL (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97: 6640–6645. doi: 10.1073/pnas.120163297
|
| [34] | Casadaban MJ, Chou J, Cohen SN (1980) In vitro gene fusions that join an enzymatically active beta-galactosidase segment to amino-terminal fragments of exogenous proteins: Escherichia coli plasmid vectors for the detection and cloning of translational initiation signals. J Bacteriol 143: 971–980. |
| [35] |
Tramonti A, De Canio M, Delany I, et al. (2006) Mechanisms of transcription activation exerted by GadX and GadW at the gadA and gadBC gene promoters of the glutamate-based acid resistance system in Escherichia coli. J Bacteriol 188: 8118–8127. doi: 10.1128/JB.01044-06
|
| [36] |
Occhialini A, Jimenez de Bagues MP, Saadeh B, et al. (2012) The glutamic acid decarboxylase system of the new species Brucella microti contributes to its acid resistance and to oral infection of mice. J Infect Dis 206: 1424–1432. doi: 10.1093/infdis/jis522
|
| [37] |
De Biase D, Tramonti A, John RA, et al. (1996) Isolation, overexpression, and biochemical characterization of the two isoforms of glutamic acid decarboxylase from Escherichia coli. Protein Expr Purif 8: 430–438. doi: 10.1006/prep.1996.0121
|
| [38] |
Capitani G, De Biase D, Aurizi C, et al. (2003) Crystal structure and functional analysis of Escherichia coli glutamate decarboxylase. Embo J 22: 4027–4037. doi: 10.1093/emboj/cdg403
|
| [39] |
Lin J, Lee IS, Frey J, et al. (1995) Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli. J Bacteriol 177: 4097–4104. doi: 10.1128/jb.177.14.4097-4104.1995
|
| [40] |
Riley M, Abe T, Arnaud MB, et al. (2006) Escherichia coli K-12: a cooperatively developed annotation snapshot-2005. Nucleic Acids Res 34: 1–9. doi: 10.1093/nar/gkj405
|
| [41] |
Altschul SF, Madden TL, Schaffer AA, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. doi: 10.1093/nar/25.17.3389
|
| [42] |
Burkovski A, Kramer R (2002) Bacterial amino acid transport proteins: occurrence, functions, and significance for biotechnological applications. Appl Microbiol Biotechnol 58: 265–274. doi: 10.1007/s00253-001-0869-4
|
| [43] |
Richard H, Foster JW (2004) Escherichia coli glutamate- and arginine-dependent acid resistance systems increase internal pH and reverse transmembrane potential. J Bacteriol 186: 6032–6041. doi: 10.1128/JB.186.18.6032-6041.2004
|
| [44] | Anderson MA, Mann MD, Evans MA, et al. (2016) The inner membrane protein YhiM is necessary for Escherichia coli growth at high temperatures and low osmolarity. Arch Microbiol. |
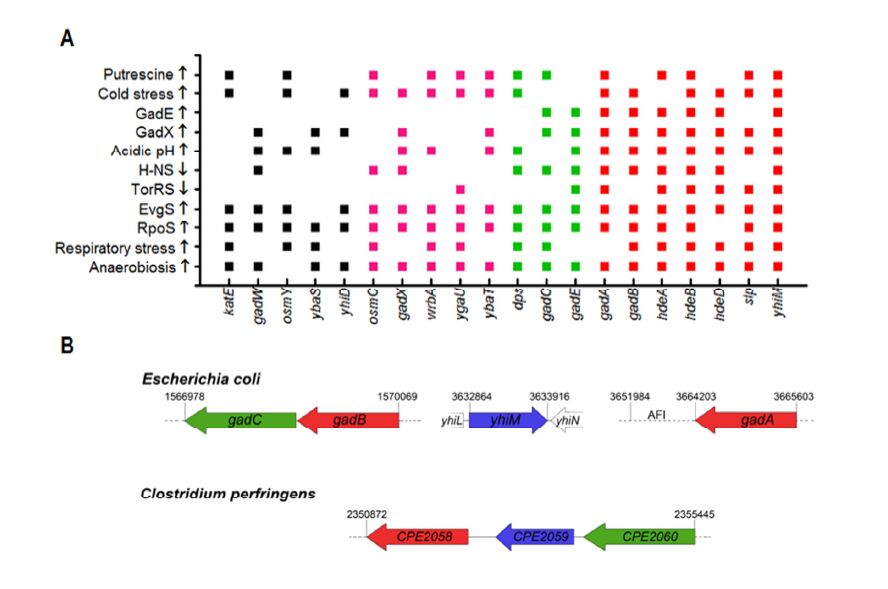

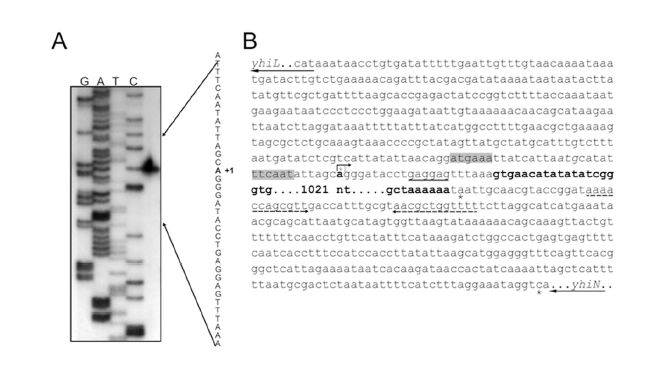
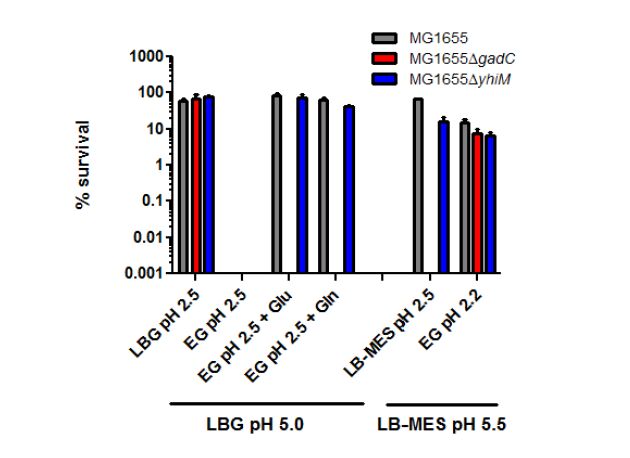
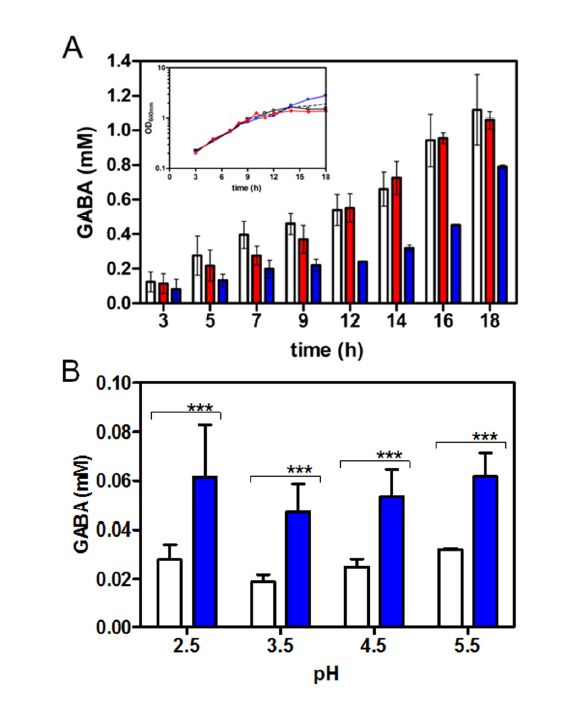
Figures(5) / Tables(4)

Angela Tramonti, Fiorenzo De Santis, Eugenia Pennacchietti, Daniela De Biase. The yhiM gene codes for an inner membrane protein involved in GABA export in Escherichia coli[J]. AIMS Microbiology, 2017, 3(1): 71-87. doi: 10.3934/microbiol.2017.1.71







 DownLoad:
DownLoad: