Citation: Merja Lusa, Jukka Lehto, Malin Bomberg. The uptake of Ni2+ and Ag+ by bacterial strains isolated from a boreal nutrient-poor bog[J]. AIMS Microbiology, 2016, 2(2): 120-137. doi: 10.3934/microbiol.2016.2.120

| [1] | Gupta V, Rastogic A, Nayak A (2010) Biosorption of nickel onto treated alga (Oedogonium hatei): Application of isotherm and kinetic models. J Colloid Interface Sci 342 (2): 533–539. |
| [2] | Rajapaksha RMCP, Tobor-Kapłon MA, Bååth E (2004) Metal Toxicity Affects Fungal and Bacterial Activities in Soil Differently. Appl Environ Microbiol 70 (5): 2966–2973. |
| [3] |
Sigee DC, Al-Rabaee RH (1986) Nickel toxicity in Pseudomonas tabaci: Single cell and bulk sample analysis of bacteria cultured at high cation levels. Protoplasma 130: 171–185. doi: 10.1007/BF01276599

|
| [4] | Dönmez G, Aksu Z, Öztürk A, et al. (1990) A comparative study on heavy metal biosorption characteristics of some algae. Process Biochem 34: 885–892. |
| [5] | Wong J, Wong Y, Tam N (2000) Nickel biosorption by two chlorella species, C. vulgaris (a commercial species) and C. miniata (a local isolate). Biores Technol 73: 133–137. |
| [6] | Aksu Z (2002) Determination of the equilibrium, kinetic and thermodynamic parameters of the batch biosorption of nickel(II) ions onto Chlorella vulgaris Process Biochem 38: 89–99. |
| [7] | Yan G, Viraraghavan T (2003) Heavy-metal removal from aqueous solution by fungus Mucor rouxii. Water Res 37: 4486–4496. |
| [8] | Aksu Z, Dönmez G (2006) Binary biosorption of cadmium(II) and nickel(II) onto dried Chlorella vulgaris: Co-ion effect on mono-component isotherm parameters. Process Biochem 41: 860–868. |
| [9] | Akhtar, Iqbal J, Iqbal M (2004) Removal and recovery of nickel(II) from aqueous solution by loofa sponge-immobilized biomass of Chlorella sorokiniana: characterization studies. J Hazard Mater 108: 85–94. |
| [10] | Grandjean P (1984) Human exposure to nickel. IARC Sci Publ 53: 469–485. |
| [11] | Von Burg R (1997) Toxicology update. J Appl Toxicol 17: 425–431. |
| [12] | Cempel M, Nikel G (2006) Nickel: A Review of Its Sources and Environmental Toxicology. Polish J of Environ Stud 15 (3): 375–382. |
| [13] | Hjerpe T, Ikonen A, Broed R (2010) Biosphere Assessment Report 2009. Posiva report 2010-03, Posiva Oy, Eurajoki, Finland. |
| [14] | Helin J, Hjerpe T, Ikonen A (2010) Review of Element Specific Data for Biosphere Assessment BSA-2009. Working Report 2010-37, Posiva Oy, Eurajoki, Finland. |
| [15] | Hjerpe T, Broed R (2010) Radionuclide Transport and Dose Assessment Modelling in Biosphere Assessment 2009. Working report 2010-79, Posiva Oy, Eurajoki, Finland. |
| [16] |
Chen C, Lin T (1998) Nickel toxicity to human term placenta: in vitro study on lipid peroxidation. J Toxicol Environ Health 54: 37–47. doi: 10.1080/009841098159015
|
| [17] | Scott-Fordsmand J (1997) Toxicity of nickel to soil organisms in Denmark. Rev Environ Contam Toxicol 148: 1–34. |
| [18] |
Bennett B (1982) Exposure of man to environmental nickel–an exposure commitment assessment. Sci Total Environ 22: 203–212. doi: 10.1016/0048-9697(82)90065-1
|
| [19] |
Pedroso M, Pinho G, Rodrigues S, et al. (2007) Mechanism of acute silver toxicity in the euryhaline copepod. Acartia tonsa Aquat Toxicol 82: 173–180. doi: 10.1016/j.aquatox.2007.02.009
|
| [20] |
Jacobson A, McBride M, Baveye P, et al. (2005) Environmental factors determining the trace-level sorption of silver and thallium to soils. Sci Total Environ 345: 191–205. doi: 10.1016/j.scitotenv.2004.10.027
|
| [21] | Smith I, Carson B (1977) Silver. Ann Arbor, MI7 Ann Arbor Science Publishers. |
| [22] | Rosenman K, Seixas N, Jacobs I (1987) Potential nephrotoxic effects of exposure to silver, Br J Ind Med 44: 267–272. |
| [23] | Das D, Das N, Mathew L (2010) Kinetics, equilibrium and thermodynamic studies on biosorption of Ag(I) from aqueous solution by macrofungus Pleurotus platypus. J Hazard Mater 184: 765–774. |
| [24] |
Jones K, Davies B, Peterson P (1986) Silver in Welsh soils: physical and chemical distribution studies. Geoderma 37: 157–74. doi: 10.1016/0016-7061(86)90028-5
|
| [25] |
Klitzke S, Metreveli G, Peters A, et al. (2015) The fate of silver nanoparticles in soil solution-Sorption of solutes and aggregation. Sci Total Environ 535: 54–60. doi: 10.1016/j.scitotenv.2014.10.108
|
| [26] | SKB (2011) Long-term safety for the final repository for spent nuclear fuel at Forsmark Main report of the SR-Site project Volume III. Errata 2011-10. SKB TR-11-01. Svensk Kärnbränslehantering AB, Stockholm. |
| [27] | Lopéz A, Lázaro N, Priego J, et al. (2010) Effect of pH on the biosorption of nickel and other heavy metals by Pseudomonas fluorescens 4F39. J Ind Microbiol Biotechnol 24: 146–151. |
| [28] |
Veglio F, Beolchini F (1997) Removal of metals by biosorption: a review. Hydrometallurgy 44: 301–316. doi: 10.1016/S0304-386X(96)00059-X
|
| [29] | Rani G, Prerna A, Seema K, et al. (2000). Microbial biosorbents: meeting challenges of heavy metal pollution in aqueous solutions, Curr Sci 8: 967–973. |
| [30] |
Tsezos M, Remoudaki E, Angelatou V (1996) A study of the effects of competing ions on the biosorption of metals. Int Biodeter Biodegrad 38: 19–29. doi: 10.1016/S0964-8305(96)00011-X
|
| [31] |
Veglio F, Beolchini F, Gasbarro A (1997) Biosorption of toxic metals: an equilibrium study using free cells of Arthrobacter sp. Process Biochem 32: 99–105. doi: 10.1016/S0032-9592(96)00047-7
|
| [32] | Paul S, Bera D, Chattopadhyay P (2006) Biosorption of lead by Bacillus cereus M 1 16 immobilized in Ca-alginate gel. J Hazard Sub Res 5: 2–6. |
| [33] |
Beveridge T (1989) The role of cellular design in bacterial metal accumulation and mineralization. Annu Rev Microbiol 43: 147–171. doi: 10.1146/annurev.mi.43.100189.001051
|
| [34] |
Volesky B (2001) Detoxification of metal-bearing effluents: biosorption for the next century. Hydrometallurgy 59: 203–216. doi: 10.1016/S0304-386X(00)00160-2
|
| [35] | Tahir A, Shehzadi R, Mateen B, et al. (2009) Biosorption of nickel (II) from effluent of electroplating industry by immobilized cells of Bacillus species, Eng Life Sci 9 (6): 462–467. |
| [36] |
Brady D, Stoll A, Duncan J (1994) Biosorption of heavy metal cations by non-viable yeast biomass. Environ Technol 15: 429–438. doi: 10.1080/09593339409385447
|
| [37] | Corder S, Reeves M (1994) Biosorption of nickel in complex aqueous waste streams by cyanobacteria. Appl Biochem Biotechnol 45/46: 847–859. |
| [38] | Volesky B (1994) Advances in biosorption of metals: selection of biomass types, FEMS Microbiol Rev 14: 291–302. |
| [39] |
Sar P, Kazy R, Asthana R, et al. (1999) Metal adsorption and desorption by lyophilized Pseudomonas aeruginosa. Int Biodeter Biodegrad 44: 101–110. doi: 10.1016/S0964-8305(99)00064-5
|
| [40] |
Kapoor A, Viraraghavan T, Cullimore P (1999) Removal of heavy metals using the fungus Aspergillus niger. Biores Technol 70: 95–104. doi: 10.1016/S0960-8524(98)00192-8
|
| [41] |
Yu Q, Matheickal J, Yin P, et al. (1999) Heavy metal uptake capacities of common marine macro algal biomass. Water Res 33: 1534–1537. doi: 10.1016/S0043-1354(98)00363-7
|
| [42] |
Abou-Shanab R, Angle J, Delorme T, et al. (2003) Rhizobacterial effects on nickel extraction from soil and uptake by Alyssum murale. New Phytologist 158: 219–224. doi: 10.1046/j.1469-8137.2003.00721.x
|
| [43] |
Mulrooney S, Hausinger R (2003) Nickel uptake and utilization by microorganisms. FEMS Microbiol Rev. 27: 239–261. doi: 10.1016/S0168-6445(03)00042-1
|
| [44] |
Zawadzka A, Crawford R., Paszczynski A (2006) Pyridine-2,6-bis(thiocarboxylic acid) produced by Pseudomonas stutzeri KC reduces and precipitates selenium and tellurium oxyanions. Appl Environ Microbiol 72: 3119–3129. doi: 10.1128/AEM.72.5.3119-3129.2006
|
| [45] | Lusa M, Lehto J, Aromaa H, et al. (2016) Uptake on radioiodide by Paenibacillus sp., Pseudomonas sp., Burkholderia sp. and Rhodococcus sp. isolated from a boreal nutrient-poor bog. J Environ Sci [In press]. |
| [46] | Mäkilä M, Grundström A (2008) Turpeen ikä ja kerrostumisnopeus Lounais-Suomen soilla. Posiva WR 2008–12. |
| [47] | Haapanen R, Aro L, Helin J, et al. (2013). Studies on Reference Mires: 1. Lastensuo and Pesänsuo in 2010–2011. Working Report 2012–102, Posiva Oy, Eurajoki, Finland. |
| [48] | Finnish meteorological institute (2015) Kuukausitilastot, available from: http://ilmatieteenlaitos.fi/kuukausitilastot |
| [49] | Tsitko I, Lusa M, Lehto J, et al. (2014) The Variation of Microbial Communities in a Depth Profile of an Acidic, Nutrient-Poor Boreal Bog in Southwestern Finland. OJE 4: 832–859. |
| [50] | Lusa M, Bomberg M, Aromaa H, et al. (2015A) Sorption of radioiodide in an acidic, nutrient-poor boreal bog: Insights into the microbial impact. J Environ Radioact143: 110–122. |
| [51] | Neidhard, F, Umbarger H (1996) Escherichia coli and Salmonella: Cellular and Molecular Biology. Vol 1 Chapter 3, Chemical Composition of Escherichia coli. ASM Press. pp. 14. |
| [52] |
Srinivasa Rao P, Kalyani S, Suresh Reddy K, et al. (2005) Comparison of Biosorption of Nickel (II) and Copper (II) Ions from Aqueous Solution by Sphaeroplea Algae and Acid Treated Sphaeroplea Alga. Sep Sci Technol 40: 3149–3165. doi: 10.1080/01496390500385350
|
| [53] |
Amini M, Younesi H, Bahramifar N (2009) Biosorption of nickel(II) from aqueous solution by Aspergillus niger: Response surface methodology and isotherm study. Chemosphere 75: 1483–1491. doi: 10.1016/j.chemosphere.2009.02.025
|
| [54] | Sujatha P, Kalarani V, Naresh Kumar B (2013) Effective Biosorption of Nickel(II) from Aqueous Solutions Using Trichoderma viride. J Chem 2013: 716098. |
| [55] | Gelagutashvili E, Ginturi E, Kuchava N, et al. (2011) Biosorption of Ag(I)-Spirulina platensis for different pH. Chem Phys arXiv:1106.0594. |
| [56] |
Gelagutashvili E (2013) Comparative Study on Heavy Metals Biosorption by Different Types of Bacteria. OJMETAL 3: 62–67. doi: 10.4236/ojmetal.2013.32A1008
|
| [57] |
Chen C, Wen D, Wang J (2014) Cellular surface characteristics of Saccharomyces cerevisiae before and after Ag(I) biosorption. Bioresour Technol 156: 380–383. doi: 10.1016/j.biortech.2014.01.065
|
| [58] |
Simmons P, Singleton I (1996) A method to increase silver biosorption by an industrial strain of Saccharomyces cerevisiae. Appl Microbiol Biotechnol 45: 278–285. doi: 10.1007/s002530050684
|
| [59] | Sun D, Li X, Zhang G (2013) Biosorption of Ag(I) from Aqueous Solution by Bacillus licheniformis Strain R08. Appl Mech Mater 295–298: 129–134. |
| [60] |
Vijayaraghavan K, Yun Y (2008) Bacterial biosorbents and biosorption. Biotechnol Adv 26: 266–291. doi: 10.1016/j.biotechadv.2008.02.002
|
| [61] | Andersen R, Chapman S, Artz R (2013) Microbial Communities in Natural and Disturbed Peatlands: A Review. Soil Biol Biochem 57: 979–994. |
| [62] |
Juottonen H, Galand P, Tuittila E, et al. (2005) Methanogen communities and Bacteria along an ecohydrological gradient in a northern raised bog complex. Environ Microbiol 7: 1547–1557. doi: 10.1111/j.1462-2920.2005.00838.x
|
| [63] |
Dedysh S, Pankratov T, Belova S, et al. (2006) Phylogenetic Analysis and In situ Identification of Bacteria Community Composition in an Acidic Sphagnum Peat Bog. Appl Environ Microbiol 72: 2110–2117. doi: 10.1128/AEM.72.3.2110-2117.2006
|
| [64] |
Sun H, Terhonen E, Koskinen K, et al. (2014) Bacterial diversity and community structure along different peat soils in boreal forest. Appl Soil Ecol 74: 37–45. doi: 10.1016/j.apsoil.2013.09.010
|
| [65] | Gadd M, White C (1993) Microbial Treatment of Metal Pollution—A Working Biotechnology, Trends Biotechnol 11: 353–359. |
| [66] |
Mishra B, Boyanov M, Bunker B, et al. (2010) High- and low-affinity binding sites for Cd on the bacterial cell walls of Bacillus subtilis and Shewanella oneidensis. Geochim. Cosmochim Acta 74: 4219–4233. doi: 10.1016/j.gca.2010.02.019
|
| [67] |
Rosen B, Liu Z (2009) Transport pathways for arsenic and selenium: A minireview. Environ Int 35: 512–515. doi: 10.1016/j.envint.2008.07.023
|
| [68] |
Turner R, Weiner J, Tylor D (1998) Selenium metabolism in Escheria coli. Biometals 11: 223–227. doi: 10.1023/A:1009290213301
|
| [69] | Lusa M, Bomberg M, Aromaa H, et al. (2015B) The microbial impact on the sorption behaviour of selenite in an acidic, nutrient-poor boreal bog. J Environ Radioact 147: 85–96. |
| [70] | Lusa M, Bomberg M, Virtanen S, et al. (2015C). Factors affecting the sorption of cesium in a nutrient-poor boreal bog. J Environ Radioact 147: 22–32. |
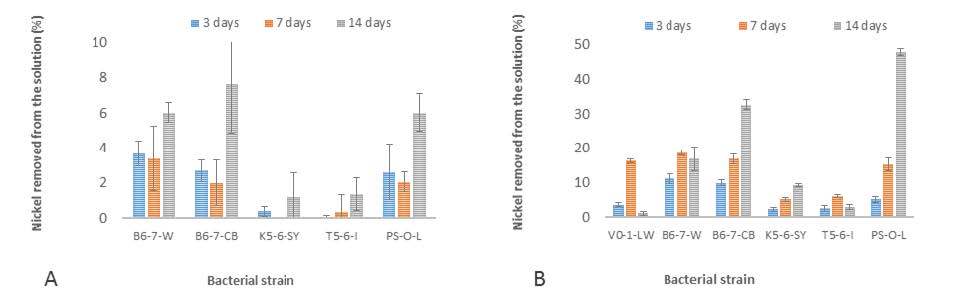
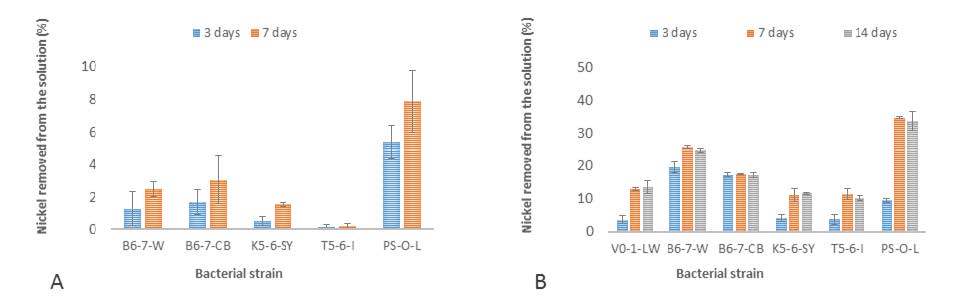
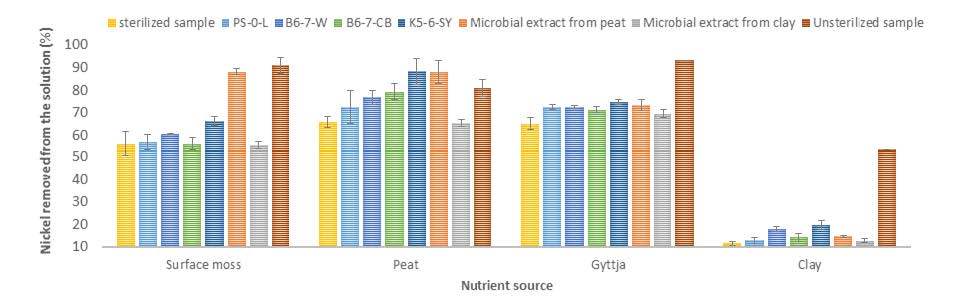
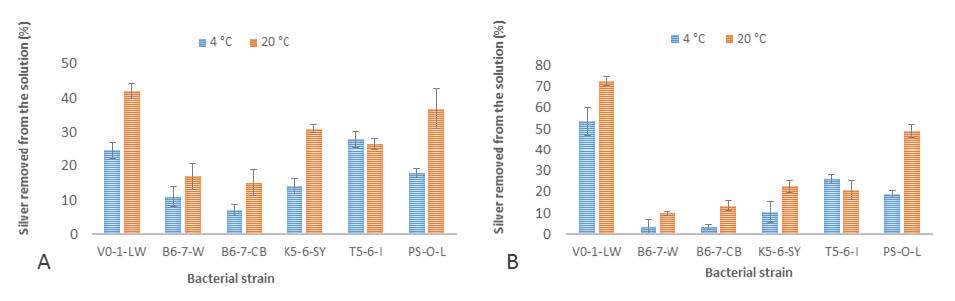
Figures(4) / Tables(2)

Merja Lusa, Jukka Lehto, Malin Bomberg. The uptake of Ni2+ and Ag+ by bacterial strains isolated from a boreal nutrient-poor bog[J]. AIMS Microbiology, 2016, 2(2): 120-137. doi: 10.3934/microbiol.2016.2.120







 DownLoad:
DownLoad: