Citation: Asha Usha Vijayakumar, Manju Sudhakaran, Leelabhai Janaki Yovel. Human basilar artery: morphology & variations[J]. AIMS Medical Science, 2020, 7(4): 278-292. doi: 10.3934/medsci.2020017

| [1] | Standring S, Ellis H, Wigley C (2005) Gray's anatomy: The anatomical basis of clinical practice Edinburgh; New York: Elsevier Churchill Livingstone. |
| [2] | Wankhede HA, Hosmani PB, Nimje DA (2014) Morphological study of the basilar artery in adult human cadavers. Int J Anat Res 2: 497-502. |
| [3] | Efendic A, Isakovic E, Delic J, et al. (2014) Vascular geometry of vertebrobasilar tree with and without aneurysm. Med Glas (Zenica) 11: 252-257. |
| [4] |
Ravensbergen J, Hillen B, Krijger JK, et al. (1996) The influence of the angle of confluence on the flow in a Vertebrobasilar junction model. J Biomech 29: 281-299. doi: 10.1016/0021-9290(95)00064-X

|
| [5] |
Tanaka M, Sakaguchi M, Miwa K, et al. (2013) Basilar artery diameter is an independent predictor of incident cardiovascular events. Arterioscler Thromb Vasc Biol 33: 2240-2244. doi: 10.1161/ATVBAHA.113.301467
|
| [6] |
Aoki J, Iguchi Y, Kimura K, et al. (2010) Diameter of the basilar artery may be associated with neurological deterioration in acute pontine infarction. Eur Neurol 63: 221-226. doi: 10.1159/000279619
|
| [7] |
Menshawi K, Gutierrez J, Mohr JP (2015) A functional perspective on embryology and anatomy of cerebral blood supply. J Stroke 17: 144-158. doi: 10.5853/jos.2015.17.2.144
|
| [8] |
van Raamt AF, van Laar PJ, Mali WP, et al. (2006) The fetal variant of the circle of Willis and its influence on the cerebral collateral circulation. Cerebrovasc Dis 22: 217-224. doi: 10.1159/000094007
|
| [9] | Romanes GJ (1986) Cunningham's Manual of Practical Anatomy Oxford University Press. |
| [10] | Padmavathi G, Niranjana Murthy KV, Rajeshwari T (2011) Study of the variations in the origin and termination of basilar artery. Anatomica Karnataka 5: 54-59. |
| [11] | Yaşargil MG (2009) Microneurosurgery: Principles, applications, and training. Practical Handbook of Neurosurgery Vienna: Springer. |
| [12] |
Fisher CM, Gore I, Okabe N, et al. (1965) Atherosclerosis of the carotid and vertebral arteries—extracranial and intracranial. J Neuropathol Exp Neurol 24: 455-476. doi: 10.1097/00005072-196507000-00007
|
| [13] |
Hong JM, Chung CS, Bang OY, et al. (2009) Vertebral artery dominance contributes to basilar artery curvature and peri-vertebrobasilar junctional infarcts. J Neurol Neurosurg Psychiatry 80: 1087-1092. doi: 10.1136/jnnp.2008.169805
|
| [14] | Mamatha H, D'souza AS, Suhani S (2012) Human cadaveric study of the morphology of the basilar artery. Singapore Med J 53: 760-763. |
| [15] | Iqbal S (2013) Vertebro-basilar variants and their basic clinical implications. Int J Med Res Health Sci 2: 799-808. |
| [16] | Wankhede HA, Hosmani PB, Nimje DA (2014) Morphological study of the basilar artery in adult human cadavers. Int J Anat Res 2: 497-502. |
| [17] |
Pai BS, Varma RG, Kulkarni RN, et al. (2007) Microsurgical anatomy of the posterior circulation. Neurology India 55: 31-41. doi: 10.4103/0028-3886.30424
|
| [18] | Glagov S, Zarins C, Giddens DP, et al. (1988) Hemodynamics and atherosclerosis. Insights and perspectives gained from studies of human arteries. Arch Pathol Lab Med 112: 1018-1031. |
| [19] |
Lee SH, Hur N, Jeong SK (2012) Geometric analysis and blood flow simulation of basilar artery. J Atheroscler Thromb 19: 397-405. doi: 10.5551/jat.10694
|
| [20] |
Passero S, Filosomi G (1998) Posterior circulation infarcts in patients with vertebrobasilar dolichoectasia. Stroke 29: 653-659. doi: 10.1161/01.STR.29.3.653
|
| [21] | Sacks JG, Lindenberg R (1970) Dolicho-ectatic intracranial arteries. Symptomatology and pathogenesis of arterial elongation and distension. Johns Hopkins Med J 125: 95-105. |
| [22] | Goldstein SJ, Sacks JG, Lee C, et al. (1983) Computed tomographic findings in cerebral artery ectasia. AJNR 4: 501-504. |
| [23] |
Saeki N, Rhoton AL (1977) Anatomy of the posterior circle of Willis. J Neurosurg 46: 564-578. doi: 10.3171/jns.1977.46.5.0563
|
| [24] | Kamath SA (1979) A study of dimensions of the basilar artery in South Indian subjects. J Anat Soc India 28: 45-64. |
| [25] |
Pico F, Labreuche J, Gourfinkel-An I, et al. (2006) Basilar artery diameter and 5-year mortality in patients with stroke. Stroke 37: 2342-2347. doi: 10.1161/01.STR.0000236058.57880.03
|
| [26] |
Black SP, Ansbacher LE (1984) Saccular aneurysm associated with segmental duplication of the basilar artery. A morphological study. J Neurosurg 6: 1005-1008. doi: 10.3171/jns.1984.61.6.1005
|
| [27] |
Finlay HM, Canham PB (1994) The layered fabric of cerebral artery fenestrations. Stroke 25: 1799-1806. doi: 10.1161/01.STR.25.9.1799
|
| [28] | Stopford JS (1916) The arteries of the pons and medulla oblongata. J Anat Physiol 50: 131-164. |

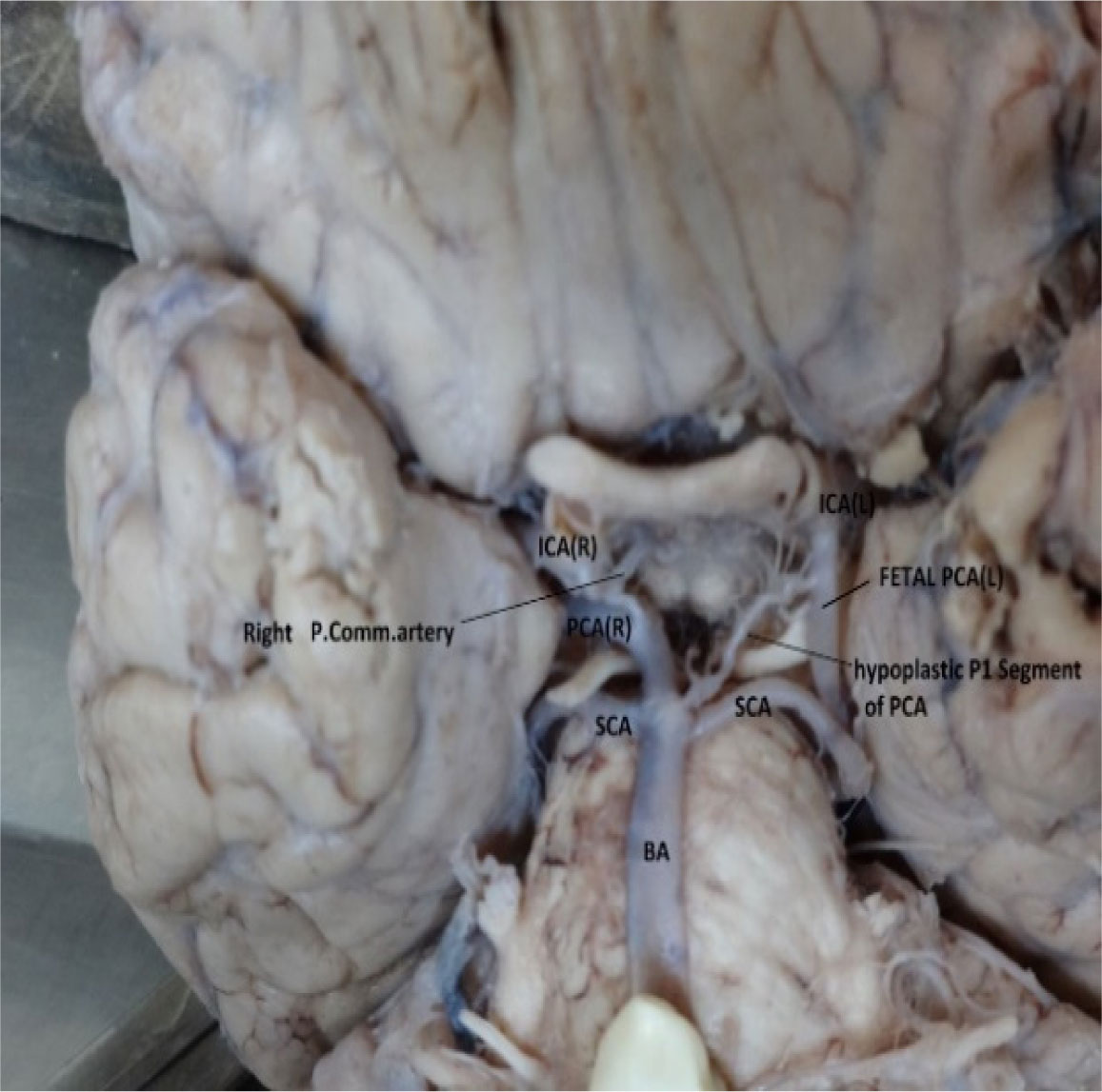
Figures(10) / Tables(5)

Asha Usha Vijayakumar, Manju Sudhakaran, Leelabhai Janaki Yovel. Human basilar artery: morphology & variations[J]. AIMS Medical Science, 2020, 7(4): 278-292. doi: 10.3934/medsci.2020017







 DownLoad:
DownLoad: