Citation: Tyler C. Smith, Besa Smith. Consistency in Physical Activity and Increase in Mental Health in Elderly over a Decade: Are We Achieving Better Population Health?[J]. AIMS Medical Science, 2016, 3(1): 147-161. doi: 10.3934/medsci.2016.1.147

| [1] | Armstrong GL, Conn LA, Pinner RW (1999) Trends in infectious disease mortality in the United States during the 20th century. JAMA 281: 61-66. |
| [2] | Hamlin C, Sheard S (1998) Revolutions in public health: 1848, and 1998? BMJ 317: 587-591. |
| [3] | Leading Causes of Death, 1900-1998. |
| [4] |
Breslow L (1999) From disease prevention to health promotion. JAMA 281: 1030-1033. doi: 10.1001/jama.281.11.1030

|
| [5] | Institute of Medicine. Committee on Quality Care in America. (2001) CROSSING THE QUALITY CHASM: A New Health System for the 21st Century. Washington, DC: Institute of Medicine. |
| [6] | Winslow CE (1920) The Untilled Fields of Public Health. Science 51: 23-33. |
| [7] | Centers for Disease C, Prevention. (2011) Ten great public health achievements--United States, 2001-2010. MMWR. Morbidity and mortality weekly report 60: 619-623. |
| [8] | Centers for Disease C, Prevention. (1999) Ten great public health achievements--United States, 1900-1999. MMWR. Morbidity and mortality weekly report 48: 241-243. |
| [9] | Arias E (2015) United States Life Tables, 2011. National vital statistics reports : from the Centers for Disease Control and Prevention, National Center for Health Statistics, National Vital Statistics System. 64: 1-63. |
| [10] | Olshansky SJ, Passaro DJ, Hershow RC, et al. (2005) A potential decline in life expectancy in the United States in the 21st century. The New England Journal of Medicine 352: 1138-1145. |
| [11] | Oeppen J, Vaupel JW (2002) Demography. Broken limits to life expectancy. Science 296: 1029-1031. |
| [12] | Medicine IIo. A Nationwide Framework for Surveillance of Cardiovascular and Chronic Lung Disease. Washington, DC2011. |
| [13] | Nelson DE, Holtzman D, Bolen J, et al. (2001) Reliability and validity of measures from the Behavioral Risk Factor Surveillance System (BRFSS). Sozial- und Praventivmedizin 46: S3-42. |
| [14] | Stein AD, Lederman RI, Shea S (1993) The Behavioral Risk Factor Surveillance System questionnaire: its reliability in a statewide sample. Am J Public Health 83: 1768-1772. |
| [15] | Li C, Balluz LS, Ford ES, et al. (2012) A comparison of prevalence estimates for selected health indicators and chronic diseases or conditions from the Behavioral Risk Factor Surveillance System, the National Health Interview Survey, and the National Health and Nutrition Examination Survey, 2007-2008. Prev Med 54: 381-387. |
| [16] | Arday DR, Tomar SL, Nelson DE, et al. (1987) State smoking prevalence estimates: a comparison of the Behavioral Risk Factor Surveillance System and current population surveys. Am J Public Health 87: 1665-1669. |
| [17] |
Hu SS, Balluz L, Battaglia MP, et al. (2011) Improving public health surveillance using a dual-frame survey of landline and cell phone numbers. Am J Epidemiol 173: 703-711. |
| [18] | A Nationwide Framework for Surveillance of Cardiovascular and Chronic Lung Diseases. Washington (DC) 2011. |
| [19] | Carlson SA, Densmore D, Fulton JE, et al. (2009) Differences in physical activity prevalence and trends from 3 U.S. surveillance systems: NHIS, NHANES, and BRFSS. J Physi Act Health 6: S18-27. |
| [20] | Aging. USDoHaHSAo. Aging statistics. http://www.aoa.acl.gov/aging_statistics/index.aspx. Accessed January 13, 2016. |
| [21] |
Nelson ME, Rejeski WJ, Blair SN, et al. (2007) Physical activity and public health in older adults: recommendation from the American College of Sports Medicine and the American Heart Association. Circulation 116: 1094-1105. doi: 10.1161/CIRCULATIONAHA.107.185650
|
| [22] | 2008 Physical Activity Guidelines for Americans. US Department of Health and Human Services; 2008. |
| [23] | Larson EB, Wang L, Bowen JD, et al. (2006) Exercise is associated with reduced risk for incident dementia among persons 65 years of age and older. An Int Med 144: 73-81. |
| [24] | DiPietro L (2001) Physical activity in aging: changes in patterns and their relationship to health and function. The journals of gerontology. Series A, Biological sciences and medical sciences. 56: 13-22. |
| [25] |
Jones DW, Peterson ED, Bonow RO, et al. (2008) Translating research into practice for healthcare providers: the American Heart Association's strategy for building healthier lives, free of cardiovascular diseases and stroke. Circulation 118: 687-696. doi: 10.1161/CIRCULATIONAHA.108.189934
|
| [26] | Clays E, Lidegaard M, De Bacquer D, et al. (2014) The combined relationship of occupational and leisure-time physical activity with all-cause mortality among men, accounting for physical fitness. Am J Epidemiol 179: 559-566. |
| [27] | Kampert JB, Blair SN, Barlow CE, et al. (1996) Physical activity, physical fitness, and all-cause and cancer mortality: a prospective study of men and women. Ann Epidemiol 6: 452-457. |
| [28] | Blair SN, Kohl HW, Paffenbarger RS, et al. (1989) Physical fitness and all-cause mortality. A prospective study of healthy men and women. JAMA 262: 2395-2401. |
| [29] | Smith TC, Wingard DL, Smith B, et al. (2007) Walking decreased risk of cardiovascular disease mortality in older adults with diabetes. J Clin Epidemiol 60: 309-317. |
| [30] | Prohaska T, Belansky E, Belza B, et al. (2006) Physical activity, public health, and aging: critical issues and research priorities. The journals of gerontology. Series B, Psychological sciences and social sciences 61: S267-273. |
| [31] | Slingerland AS, van Lenthe FJ, Jukema JW, et al. (2007) Aging, retirement, and changes in physical activity: prospective cohort findings from the GLOBE study. Am J Epidemiol 165 :1356-1363. |
| [32] | Littman A, Jacobson IG, Boyko EJ, et al. (2015) Changes in Meeting Vigorous Physical Activity Guidelines After Discharge From the Military. J Physi Act Health 12: 666-674. |
| [33] | Reilly T, Waterhouse J, Atkinson G. (1997) Aging, rhythms of physical performance, and adjustment to changes in the sleep-activity cycle. Occu Envir Med 54: 812-816. |
| [34] | Reuter I (2012) Aging, physical activity, and disease prevention. J Aging Res 2012: 373294. |
| [35] | Martin LG, Schoeni RF, Andreski PM (2010) Trends in health of older adults in the United States: past, present, future. Demography 47: S17-40. |
| [36] |
Seeman TE, Merkin SS, Crimmins EM, et al. (2010) Disability trends among older Americans: National Health And Nutrition Examination Surveys, 1988-1994 and 1999-2004. Am J Public Health 100: 100-107. doi: 10.2105/AJPH.2008.157388
|
| [37] | Lim K, Taylor L (2005) Factors associated with physical activity among older people—a population-based study. Prev Med 40: 33-40. |
| [38] | Florindo AA, Guimaraes VV, Cesar CL, et al. (2009) Epidemiology of leisure, transportation, occupational, and household physical activity: prevalence and associated factors. J Physi Act Health 6: 625-632. |
| [39] | Todt K, Skargren E, Jakobsson P, et al. (2015) Factors associated with low physical activity in patients with chronic obstructive pulmonary disease: a cross-sectional study. Scandinavian J Caring Scie 29: 697-707. |
| [40] | Healthy People 2020 Washington, DC: U.S. Department of Health and Human Services, Office of Disease Prevention and Health Promotion. |
| [41] | Piane GM, Smith TC (2014) Building an evidence base for the co-occurrence of chronic disease and psychiatric distress and impairment. Prevent Chronic Dis11: E188. |
| [42] |
Kessler RC, Chiu WT, Demler O, et al. (2005) Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. Arch General Psychiatry 62: 617-627. |
| [43] | Kessler RC, Berglund P, Demler O, et al. (2005) Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch General Psychiatry 62: 593-602. |
| [44] | Karel MJ, Gatz M, Smyer MA (2012) Aging and mental health in the decade ahead: what psychologists need to know. The Am Psycholo 67: 184-198. |
| [45] | Institute of Medicine (IOM). The Mental Health and Substance Use Workforce for Older Adults: In Whose Hands? 2012. |
| [46] | U.S. Department of Health and Human Services. Healthy People 2010: Understanding and Improving Health. 2nd ed. Vol 2005. Washington, DC: U.S. Government Printing Office; November 2000. |
| [47] | Koh HK, Piotrowski JJ, Kumanyika S, et al. (2011) Healthy people: a 2020 vision for the social determinants approach. Health education & behavior: the official publication of the Society for Public Health Education 38: 551-557. |
| [48] | Koh HK (2010) A 2020 vision for healthy people. Eng J Med 362: 1653-1656. |
| [49] | Checkoway H, Pearce N, Kriebel D (2007) Selecting appropriate study designs to address specific research questions in occupational epidemiology. Occu Envir Med 64: 633-638. |
| [50] | Pearce N (2012) Classification of epidemiological study designs. Int J Epide 41: 393-397. |
| [51] | Rothman K, Greenland S (1998) Modern Epidemiology. Second ed. Philadelphia, PA: Lippincott-Raven. |
| [52] |
Bowling A (2005) Mode of questionnaire administration can have serious effects on data quality. J Pub Health 27: 281-291. doi: 10.1093/pubmed/fdi031
|
| [53] | Krebs NF, Himes JH, Jacobson D, et al. (2007) Assessment of child and adolescent overweight and obesity. Pediatrics 120: S193-228. |
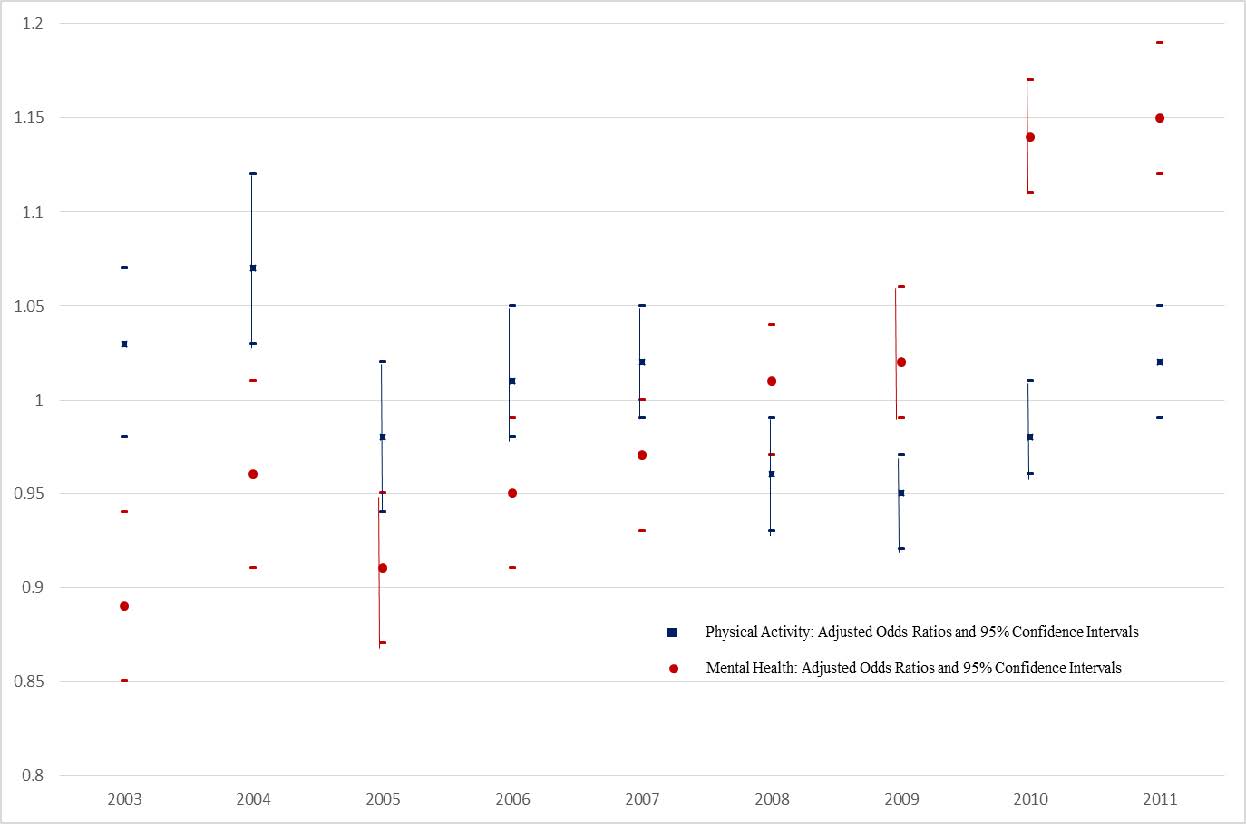
Figures(1) / Tables(2)

Tyler C. Smith, Besa Smith. Consistency in Physical Activity and Increase in Mental Health in Elderly over a Decade: Are We Achieving Better Population Health?[J]. AIMS Medical Science, 2016, 3(1): 147-161. doi: 10.3934/medsci.2016.1.147







 DownLoad:
DownLoad: