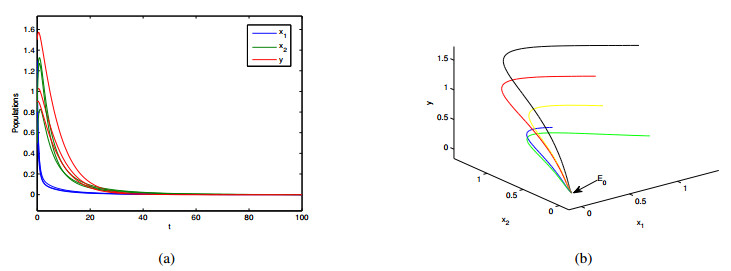
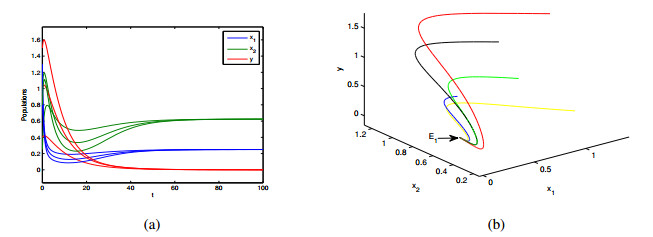
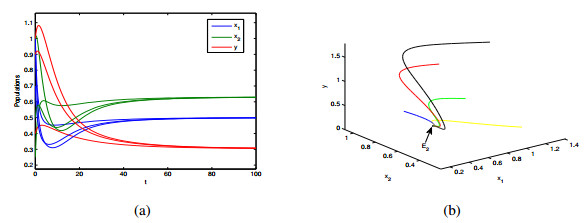
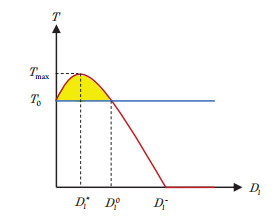
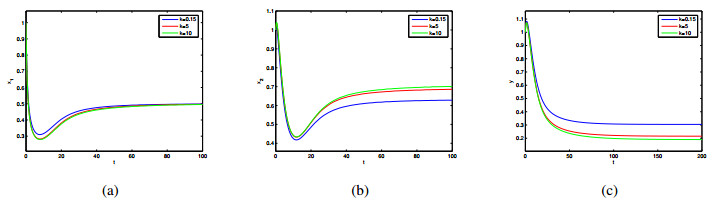
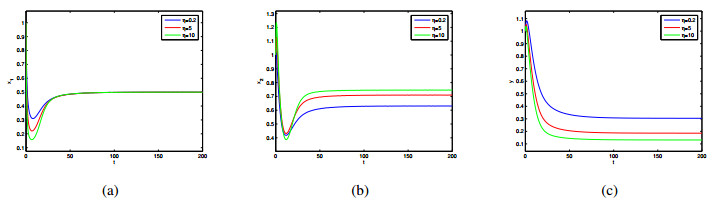
In this paper, a patchy model in which the migration is induced by the fear effect on the predator was investigated. By applying dynamical theory, the complete study on persistence of the system and the local/global stability of equilibria were discussed. Choosing the diffusion coefficient $ D_1 $ as the bifurcation parameter, transcritical bifurcation occurring near the trivial equilibrium was demonstrated. We concluded that low dispersal is favorable for the coexistence of both species, but large dispersal leads to the extinction of species. There is an optimal diffusion coefficient to make the density of the prey population reach its maximum. In addition, the level of fear effect $ k $ and the maximum fear cost $ \eta $ are beneficial to the total population density of prey.
Citation: Jin Zhong, Yue Xia, Lijuan Chen, Fengde Chen. Dynamical analysis of a predator-prey system with fear-induced dispersal between patches[J]. Mathematical Biosciences and Engineering, 2025, 22(5): 1159-1184. doi: 10.3934/mbe.2025042
In this paper, a patchy model in which the migration is induced by the fear effect on the predator was investigated. By applying dynamical theory, the complete study on persistence of the system and the local/global stability of equilibria were discussed. Choosing the diffusion coefficient $ D_1 $ as the bifurcation parameter, transcritical bifurcation occurring near the trivial equilibrium was demonstrated. We concluded that low dispersal is favorable for the coexistence of both species, but large dispersal leads to the extinction of species. There is an optimal diffusion coefficient to make the density of the prey population reach its maximum. In addition, the level of fear effect $ k $ and the maximum fear cost $ \eta $ are beneficial to the total population density of prey.

| [1] |
H. Andren, Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review, Oikos, 71 (1994), 355–366. https://doi.org/10.2307/3545823 doi: 10.2307/3545823

|
| [2] |
J. Tewksbury, D. Levey, N. Haddad, S. Sargent, J. Orrock, A. Weldon, et al., Corridors affect plants, animals, and their interactions in fragmented landscapes, Proceed. Nat. Aca. Sci., 99 (2002), 12923–12926. https://doi.org/10.1073/pnas.202242699 doi: 10.1073/pnas.202242699
|
| [3] |
F. Gilbert, A. Gonzalez, I. Evans-Freke, Corridors maintain species richness in the fragmented landscapes of a microecosystem, Proceed. Royal Soc. London. Ser. B Biol. Sci., 265 (1998), 577–582. https://doi.org/10.1098/rspb.1998.0333 doi: 10.1098/rspb.1998.0333
|
| [4] |
Y. Samia, F. Lutscher, A. Hastings, Connectivity, passability and heterogeneity interact to determine fish population persistence in river networks, J. R. Soc. Interface., 12 (2015), 20150435. https://doi.org/10.1098/rsif.2015.0435 doi: 10.1098/rsif.2015.0435
|
| [5] |
H. Freedman, P. Waltman, Mathematical models of population interactions with dispersal. I: Stability of two habitats with and without a predator, SIAM J. Appl. Math., 32 (1977), 631–648. https://doi.org/10.1137/0132052 doi: 10.1137/0132052
|
| [6] |
R. Holt, Population dynamics in two-patch environments: some anomalous consequences of an optimal habitat distribution, Theor. Popul. Biol., 28 (1985), 181–208. https://doi.org/10.1016/0040-5809(85)90027-9 doi: 10.1016/0040-5809(85)90027-9
|
| [7] |
H. Wu, Y. Wang, Dynamics of competitive systems with diffusion between source-sink patches, Bull. Math. Biol., 83 (2021), 49. https://doi.org/10.1007/s11071-021-06635-5 doi: 10.1007/s11071-021-06635-5
|
| [8] |
H. Wu, Y. Wang, Y. Li, D. DeAngelis, Dispersal asymmetry in a two-patch system with source–sink populations, Theor. Popul. Biol., 131 (2020), 54–65. https://doi.org/10.1016/j.tpb.2019.11.004 doi: 10.1016/j.tpb.2019.11.004
|
| [9] |
Y. Wang, H. Wu, Y. He, Z. Wang, K. Hu, Population abundance of two-patch competitive systems with asymmetric dispersal, J. Math. Biol., 81 (2020), 315–341. https://doi.org/10.1007/s00285-020-01511-z doi: 10.1007/s00285-020-01511-z
|
| [10] |
J. Ban, Y. Wang, H. Wu, Dynamics of predator-prey systems with prey's dispersal between patches, Indian. J. Pure Appl. Math., 53 (2022), 550–569. https://doi.org/10.1007/s13226-021-00117-5 doi: 10.1007/s13226-021-00117-5
|
| [11] |
Y. Xia, X. Huang, F. Chen, L. Chen, Stability and bifurcation analysis of a predator-prey system with multiple anti-predator behaviors, J. Biol. Syst., 32 (2024), 889-919. https://doi.org/10.1142/S021833902450030X doi: 10.1142/S021833902450030X
|
| [12] |
Y. Xia, L. J. Chen, V. Srivastava, R. Parshad, Stability and bifurcation analysis of a two-patch model with Allee effect and dispersal, Math. Biosci. Eng., 20 (2023), 1179–1197. https://doi.org/10.48550/arXiv.2310.10558 doi: 10.48550/arXiv.2310.10558
|
| [13] |
Y. Kang, S. K. Sasmal, K. Messan, A two-patch prey-predator model with predator dispersal driven by the predation strength, Math. Biosci. Eng., 14 (2017), 843–880. https://doi.org/10.3934/mbe.2017046 doi: 10.3934/mbe.2017046
|
| [14] |
A. Mai, G. Sun, F. Zhang, L. Wang, The joint impacts of dispersal delay and dispersal patterns on the stability of predator-prey metacommunities, J. Theor. Biol., 462 (2019), 455–465. https://doi.org/10.1016/j.jtbi.2018.11.035 doi: 10.1016/j.jtbi.2018.11.035
|
| [15] |
S. Saha, G. Samanta, Influence of dispersal and strong Allee effect on a two-patch predator-prey model, Int. J. Dynam. Control, 7 (2019), 1321–1349. https://doi.org/10.1007/s40435-018-0490-3 doi: 10.1007/s40435-018-0490-3
|
| [16] |
S. Biswas, D. Pal, P. Santra, E. Bonyah, G. Mahapatra, Dynamics of a three-patch prey-predator system with the impact of dispersal speed incorporating strong Allee effect on double prey, Discrete Dyn. Nat. Soc., 2022 (2022), 7919952. https://doi.org/10.1155/2022/7919952 doi: 10.1155/2022/7919952
|
| [17] |
L. Chen, T. Liu, F. Chen, Stability and bifurcation in a two-patch model with additive Allee effect, AIMS Math., 7 (2022), 536–551. https://doi.org/10.3934/math.2022034 doi: 10.3934/math.2022034
|
| [18] |
D. Luo, Q. Wang, L. Chen, Nonconstant steady states and pattern formations of generalized 1D crossdiffusion systems with prey-taxis, Stud. Appl. Math., 150 (2023), 899–958. https://doi.org/10.1111/sapm.12560 doi: 10.1111/sapm.12560
|
| [19] |
D. Luo, Q. Wang, Global bifurcation for a Beddington-DeAngelis and Tanner predator-prey reaction-diffusion system with prey-taxis, Math. Methods Appl. Sci., 47 (2024), 1711–1727. https://doi.org/10.1002/mma.9718 doi: 10.1002/mma.9718
|
| [20] |
D. Luo, Q. Wang, Spatio-temporal patterns and global bifurcation of a nonlinear cross-diffusion predator-prey model with prey-taxis and double Beddington-DeAngelis functional responses, Nonlinear Anal. Real World Appl., 79 (2024), 104133. https://doi.org/10.1016/j.nonrwa.2024.104133 doi: 10.1016/j.nonrwa.2024.104133
|
| [21] |
S. Creel, D. Christianson, Relationships between direct predation and risk effects, Trends Ecol. Evol., 23 (2008), 194–201. https://doi.org/10.1016/j.tree.2007.12.004 doi: 10.1016/j.tree.2007.12.004
|
| [22] |
W. Cresswell, Predation in bird populations, J. Ornithol., 152 (2011), 251–263. https://doi.org/10.1007/s10336-010-0638-1 doi: 10.1007/s10336-010-0638-1
|
| [23] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator–prey interactions, J. Math. Biol., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [24] |
T. Liu, L. Chen, F. Chen, Z. Li, Stability analysis of a Leslie-Gower model with strong Allee effect on prey and fear effect on predator, Int. J. Bifurcation Chaos, 32 (2022), 2250082. https://doi.org/10.1142/S0218127422500821 doi: 10.1142/S0218127422500821
|
| [25] |
T. Liu, L. Chen, F. Chen, Z. Li, Dynamics of a Leslie-Gower Model with weak Allee effect on prey and fear effect on predator, Int. J. Bifurcation Chaos, 33 (2023), 2350008. https://doi.org/10.1142/S0218127423500086 doi: 10.1142/S0218127423500086
|
| [26] |
T. Li, Q. Wang, Turing patterns in a predator–prey reaction–diffusion model with seasonality and fear effect, J. Nonlinear Sci., 33 (2023), 86. https://doi.org/10.1007/s00332-023-09938-6 doi: 10.1007/s00332-023-09938-6
|
| [27] |
Q. Cao, G. Chen, W. Yang, The impact of fear effect on the dynamics of a delayed predator-prey model with stage structure, Int. J. Biomath., 16 (2023), 2250139. https://doi.org/10.1142/S179352452250139X doi: 10.1142/S179352452250139X
|
| [28] |
X. Liu, S. Liu, Dynamics of a predator-prey system with inducible defense and disease in the prey, Nonlinear Anal. Real World Appl., 71 (2023), 103802. https://doi.org/10.1016/j.nonrwa.2022.103802 doi: 10.1016/j.nonrwa.2022.103802
|
| [29] |
K. Sarkar, S. Khajanchi, Impact of fear effect on the growth of prey in a predator-prey interaction model, Ecol. Complexity, 42 (2020), 100826. https://doi.org/10.1016/j.ecocom.2020.100826 doi: 10.1016/j.ecocom.2020.100826
|
| [30] |
Y. Dong, D. Wu, C. Shen, L. Ye, Influence of fear effect and predator-taxis sensitivity on dynamical behavior of a predator–prey model, Z. Angew. Math. Phys., 73 (2022), 25. https://doi.org/10.1007/s00033-021-01659-8 doi: 10.1007/s00033-021-01659-8
|
| [31] |
X. Meng, L. Chen, F. Chen, Dynamics of a predator-prey system with asymmetric dispersal and fear effect, Symmetry, 17 (2025), 329. https://doi.org/10.3390/sym17030329 doi: 10.3390/sym17030329
|
| [32] | H. Smith, H. Thieme, Dynamical Systems and Population Persistence, American Mathematical Soc, 2011. https://doi.org/10.1007/978-1-4684-0392-3 |
| [33] | Z. Zhang, T. Ding, W. Huang, Z. Dong, Qualitative Theory of Differential Equation, Science Press, Beijing, China, 1992. |
| [34] |
M. Li, J. Muldowney, Global stability for the SEIR model in epidemiology, Math. Biosci., 125 (1995), 155–164. https://doi.org/10.1016/0025-5564(95)92756-5 doi: 10.1016/0025-5564(95)92756-5
|
| [35] |
M. Li, J. Muldowney, Uber die lage der integralkurven gewonlicher differantialgleichungen, Proc. Phys. Math. Sot. Japan, 24 (1942), 551–559. https://doi.org/10.11429/ppmsj1919.24.0_551 doi: 10.11429/ppmsj1919.24.0_551
|
| [36] | L. Perko, Differential Equations and Dynamical Systems, Springer, New York, 1996. https://doi.org/10.1007/978-1-4684-0392-3 |
| [37] |
Y. Li, L. Liu, W. Catharine, On inverse problems in multi-population aggregation models, J. Differ. Equations, 414 (2025), 94–124. https://doi.org/10.1016/j.jde.2024.08.075 doi: 10.1016/j.jde.2024.08.075
|
| [38] |
Y. Li, L. Liu, W. Catharine, On inverse problems in predator-prey models, J. Differ. Equations, 397 (2024), 349–376. https://doi.org/10.1016/j.jde.2024.04.009 doi: 10.1016/j.jde.2024.04.009
|
| [39] |
M. Ding, L. Liu, G. Zheng, On inverse problems for several coupled PDE systems arising in mathematical biology, J. Math. Biol., 87 (2023), 86. https://doi.org/10.1007/s00285-023-02021-4 doi: 10.1007/s00285-023-02021-4
|
| [40] |
L. Liu, W. Catharine, Determining a parabolic system by boundary observation of its non-negative solutions with biological applications, Inverse Probl., 40 (2024), 025009. https://doi.org/10.1088/1361-6420/ad149f doi: 10.1088/1361-6420/ad149f
|
| [41] |
L. Liu, W. Catharine, Determining state space anomalies in mean field games, Nonlinearity, 38 (2025), 025010. https://doi.org/10.1088/1361-6544/ada67d doi: 10.1088/1361-6544/ada67d
|
| [42] |
M. Ding, L. Liu, G. Zheng, Determining a stationary mean field game system from full/partial boundary measurement, SIAM J. Math. Anal., 57 (2025), 661–681. https://doi.org/10.1137/23M1594327 doi: 10.1137/23M1594327
|
Figures(6) / Tables(1)

Jin Zhong, Yue Xia, Lijuan Chen, Fengde Chen. Dynamical analysis of a predator-prey system with fear-induced dispersal between patches[J]. Mathematical Biosciences and Engineering, 2025, 22(5): 1159-1184. doi: 10.3934/mbe.2025042







 DownLoad:
DownLoad: