Citation: Akhil Kumar Srivastav, Pankaj Kumar Tiwari, Prashant K Srivastava, Mini Ghosh, Yun Kang. A mathematical model for the impacts of face mask, hospitalization and quarantine on the dynamics of COVID-19 in India: deterministic vs. stochastic[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 182-213. doi: 10.3934/mbe.2021010

| [1] | N. M. Ferguson, D. Laydon, G. Nedjati-Gilani, N. Imai, K. Ainslie, M. Baguelin, et al., Impact of non-pharmaceutical interventions (NPIs) to reduce COVID-19 mortality and healthcare demand, London: Imperial College COVID-19 Response Team, March 16, 2020. |
| [2] |
M. McKee, D. Stuckler, If the world fails to protect the economy, COVID-19 will damage health not just now but also in the future, Nat. Med., 26 (2020), 640-642. doi: 10.1038/s41591-020-0863-y

|
| [3] |
M. Nicola, Z. Alsafi, C. Sohrabi, A. Kerwan, A. Al-Jabir, C. Iosifidis, et al., The socio-economic implications of the coronavirus pandemic (COVID-19): A review, Int. J. Surg., 78 (2020), 185-193. doi: 10.1016/j.ijsu.2020.04.018
|
| [4] |
N. van Doremalen, T. Bushmaker, D. H. Morris, M. G. Holbrook, A. Gamble, B. N. Williamson, et al., Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1, N. Engl. J. Med., 382 (2020), 1564-1567. doi: 10.1056/NEJMc2004973
|
| [5] | World Health Organization, Coronavirus disease 2019 (COVID-19). WHO situation report - 73, 2020. Avaiable from: https://www.who.int/docs/default-source/coronaviruse/situation-reports/20200402-sitrep-73-covid-19.pdf. |
| [6] | X. Liu, S. Zhang, COVID-19: face masks and human-to-human transmission. Influenza Other Respir Viruses, 2020. Avaiable from: http://doi.wiley.com/10.1111/irv.12740. |
| [7] |
C. J. Noakes, C. B. Beggs, P. A. Sleigh, K. G. Kerr, Modelling the transmission of airborne infections in enclosed spaces, Epidemiol. Infect., 134 (2006), 1082-1091. doi: 10.1017/S0950268806005875
|
| [8] | C. J. Noakes, P. A. Sleigh, Mathematical models for assessing the role of airflow on the risk of airborne infection in hospital wards, J. R. Soc. Interface, 6 (2009), S791-S800. |
| [9] |
R. Chetty, M. Stepner, S. Abraham, S. Lin, B. Scuderi, N. Turner, et al., The association between income and life expectancy in the United States, 2001-2014, JAMA, 315 (2016), 1750. doi: 10.1001/jama.2016.4226
|
| [10] |
R. O. Stutt, R. Retkute, M. Bradley, C. A. Gilligan, J. Colvin, A modelling framework to assess the likely effectiveness of facemasks in combination with 'lock-down'in managing the COVID-19 pandemic, Proc. R. Soc. A, 476 (2020), 20200376. doi: 10.1098/rspa.2020.0376
|
| [11] |
D. K. Chu, E. A. Akl, S. Duda, K. Solo, S. Yaacoub, H. J. Schunemann, et al., Physical distancing, face masks, and eye protection to prevent person-to-person transmission of SARS-CoV-2 and COVID-19: a systematic review and meta-analysis, The Lancet, 395 (2020), 1973-1987. doi: 10.1016/S0140-6736(20)31142-9
|
| [12] |
C. N. Ngonghala, E. Iboi, S. Eikenberry, M. Scotch, C. R. MacIntyre, M. H. Bonds, et al., Mathematical assessment of the impact of non-pharmaceutical interventions on curtailing the 2019 novel Coronavirus, Math. Biosci., 325 (2020), 108364. doi: 10.1016/j.mbs.2020.108364
|
| [13] | S. E. Eikenberry, M. Mancuso, E. Iboi, T. Phan, K. Eikenberry, Y. Kuang, et al., To mask or not to mask: Modeling the potential for face mask use by the general public to curtail the COVID-19 pandemic, Infect. Dis. Model., 5 (2020), 293-308. |
| [14] | E. A. Iboi, O. O. Sharomi, C. N. Ngonghala, A. B. Gumel, Mathematical modeling and analysis of COVID-19 pandemic in Nigeria, Math. Biosci. Eng., 17 (2020), 7192-7220. |
| [15] |
D. Okuonghae, A. Omame, Analysis of a mathematical model for COVID-19 population dynamics in Lagos, Nigeria, Chaos Soliton. Fract., 139 (2020), 110032. doi: 10.1016/j.chaos.2020.110032
|
| [16] |
B. J. Cowling, K. H. Chan, V. J. Fang, C. K. Cheng, R. O. Fung, W. Wai, et al., Facemasks and hand hygiene to prevent influenza transmission in households: a cluster randomized trial, Ann. Intern. Med., 151 (2009), 437-446. doi: 10.7326/0003-4819-151-7-200910060-00142
|
| [17] |
M. Lipsitch, T. Cohen, B. Cooper, J. M. Robins, S. Ma, L. James, et al., Transmission dynamics and control of severe acute respiratory syndrome, Science, 300 (2003), 1966-1970. doi: 10.1126/science.1086616
|
| [18] |
S. Zhao, L. Stone, D. Gao, S. S. Musa, M. K. Chong, D. He, et al., Imitation dynamics in the mitigation of the novel coronavirus disease (COVID-19) outbreak in Wuhan, China from 2019 to 2020, Ann. Transl. Med., 8 (2020), 448. doi: 10.21037/atm.2020.03.168
|
| [19] |
S. H. A. Khoshnaw, R. H. Salih, S. Sulaimany, Mathematical modeling for coronavirus disease (COVID-19) in predicting future behaviours and sensitivity analysis, Math. Model. Nat. Phenom., 15 (2020), 1-13. doi: 10.1051/mmnp/2019006
|
| [20] | M. Serhani, H. Labbardi, Mathematical modeling of COVID-19 spreading with asymptomatic infected and interacting peoples, J. Appl. Math. Comput., 17 (2020), 1-20. |
| [21] |
C. Huang, Y. Wang, X. Li, L. Ren, J. Zhao, Y. Hu, et al., Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China, Lancet, 395 (2020), 497-506. doi: 10.1016/S0140-6736(20)30183-5
|
| [22] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. doi: 10.1016/S0025-5564(02)00108-6
|
| [23] | COVID 2019, India, available from: https://www.covid19india.org/. |
| [24] |
L. F. White, M. Pagano, A likelihood-based method for real-time estimation of the serial interval and reproductive number of an epidemic, Stat. Med., 27 (2008), 2999-3016. doi: 10.1002/sim.3136
|
| [25] |
A. Davies, K. A. Thompson, K. Giri, G. Kafatos, J. Walker, A. Bennett, Testing the efficacy of homemade masks: would they protect in an influenza pandemic?, Disaster Med. Public Health Prep., 7 (2013), 413-418. doi: 10.1017/dmp.2013.43
|
| [26] |
C. N. Ngonghala, E. Iboi, S. Eikenberry, M. Scotch, C. R. MacIntyre, M. H. Bonds, et al., Mathematical assessment of the impact of non-pharmaceutical interventions on curtailing the 2019 novel coronavirus, Math. Biosci., 325 (2020), 108364. doi: 10.1016/j.mbs.2020.108364
|
| [27] |
S. A. Lauer, K. H. Grantz, Q. Bi, F. K. Jones, Q. Zheng, H. R. Meredith, et al., The incubation period of coronavirus disease 2019 (COVID-19) from publicly reported confirmed cases: estimation and application, Ann. Intern. Med., 172 (2020), 577-582. doi: 10.7326/M20-0504
|
| [28] |
R. Li, S. Pei, B. Chen, Y. Song, T. Zhang, W. Yang, et al., Substantial undocumented infection facilitates the rapid dissemination of novel coronavirus (SARS-CoV2), Science, 368 (2020), 489-493. doi: 10.1126/science.abb3221
|
| [29] |
F. Zhou, T. Yu, R. Du, G. Fan, Y. Liu, Z. Liu, et al., Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study, Lancet, 395 (2020), 1054-1062. doi: 10.1016/S0140-6736(20)30566-3
|
| [30] |
B. Tang, X. Wang, Q. Li, N. L. Bragazzi, S. Tang, Y. Xiao, et al., Estimation of the transmission risk of the 2019-nCoV and its implication for public health interventions, J. Clin. Med., 9 (2020), 462. doi: 10.3390/jcm9020462
|
| [31] |
D. Aldila, M. Z. Ndii, B. M. Samiadji, Optimal control on COVID-19 eradication program in Indonesia under the effect of community awareness, Math. Biosci. Eng., 17 (2020), 6355-6389. doi: 10.3934/mbe.2020335
|
| [32] |
D. He, S. Zhao, X. Xu, Q. Lin, Z. Zhuang, P. Cao, et al., Low dispersion in the infectiousness of COVID-19 cases implies difficulty in control, BMC Public Health, 20 (2020), 1558. doi: 10.1186/s12889-020-09624-2
|
| [33] |
I. Ghosh, P. K. Tiwari, J. Chattopadhyay, Effect of active case finding on dengue control: Implications from a mathematical model, J. Theor. Biol., 464 (2019), 50-62. doi: 10.1016/j.jtbi.2018.12.027
|
| [34] |
S. M. Blower, H. Dowlatabadi, Sensitivity and uncertainty analysis of complex models of disease transmission: an HIV model, as an example, Int. Stat. Rev., 62 (1994), 229-243. doi: 10.2307/1403510
|
| [35] |
S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178-196. doi: 10.1016/j.jtbi.2008.04.011
|
| [36] |
Y. Yuan, L. J. S. Allen, Stochastic models for virus and immune system dynamics, Math. Biosci., 234 (2011), 84-94. doi: 10.1016/j.mbs.2011.08.007
|
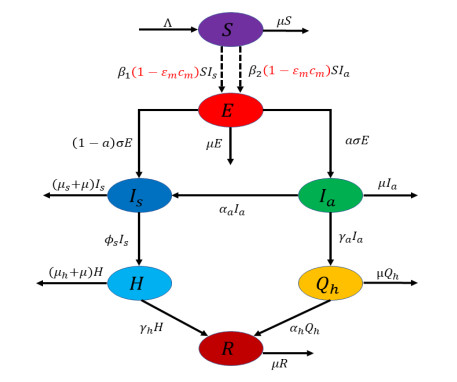
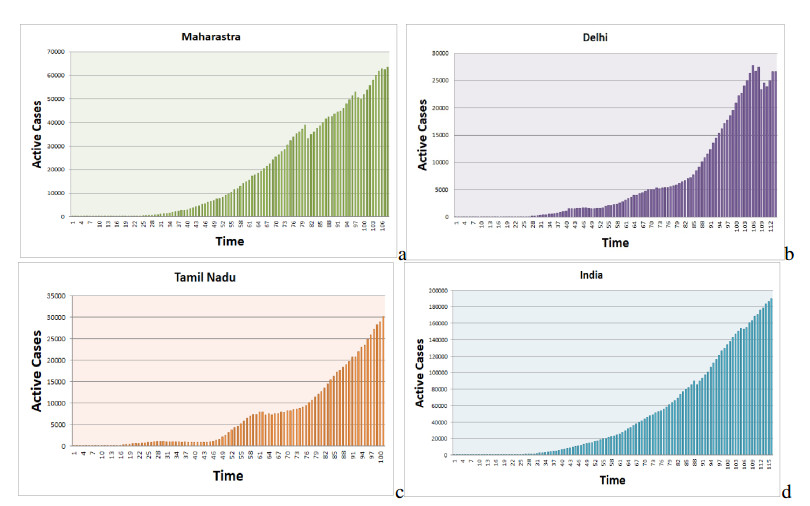
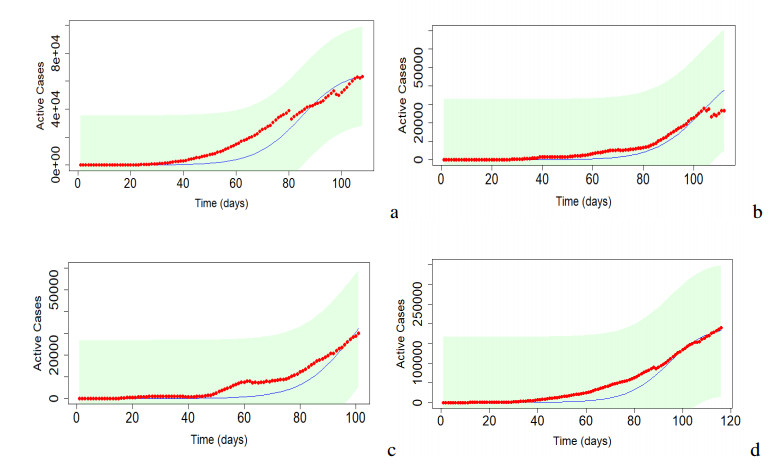
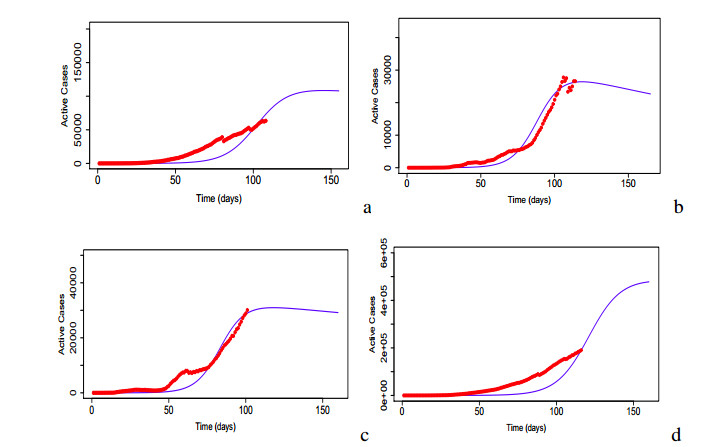
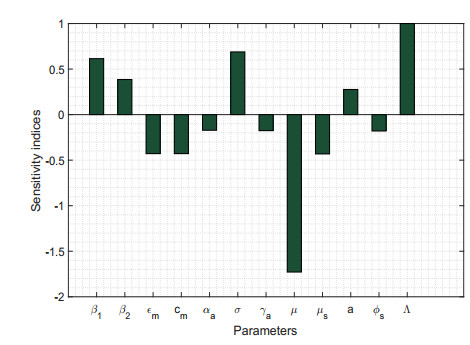
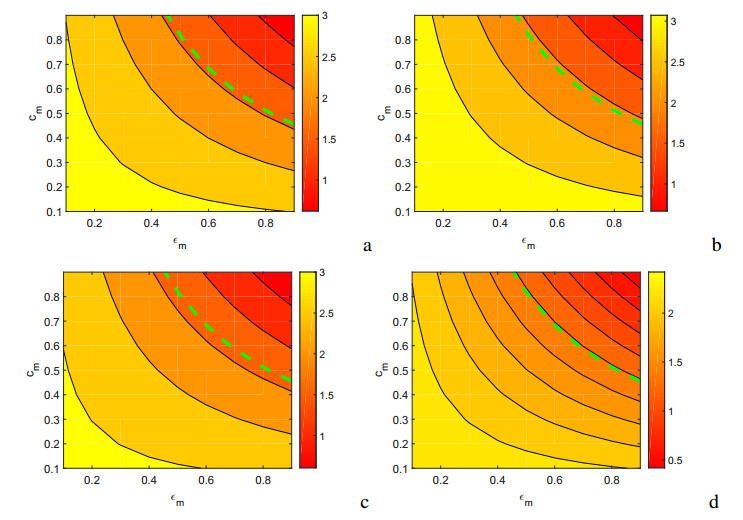
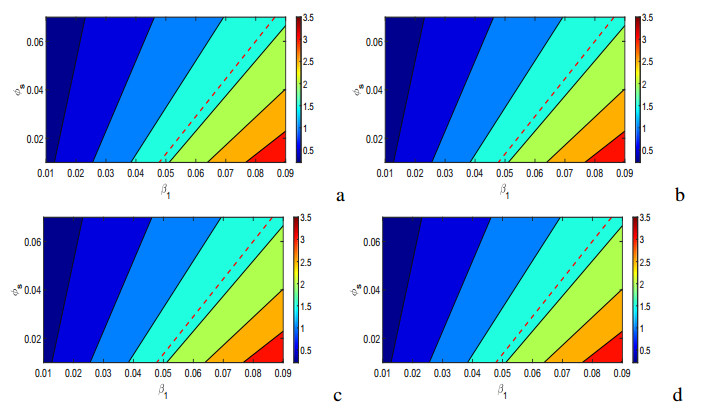
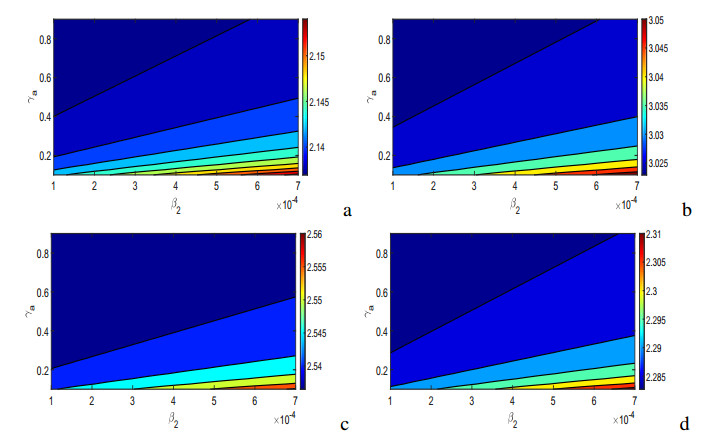
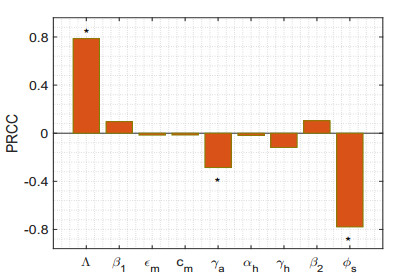
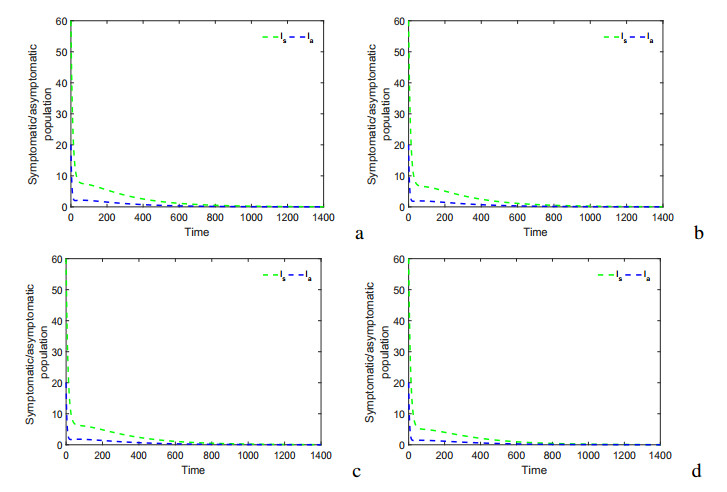
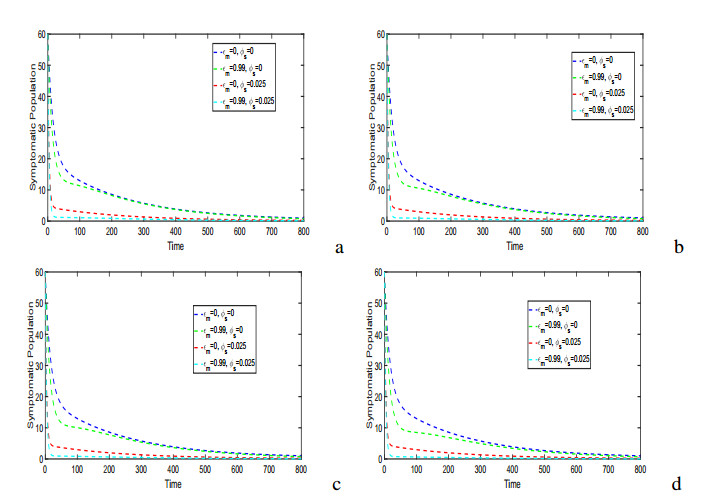
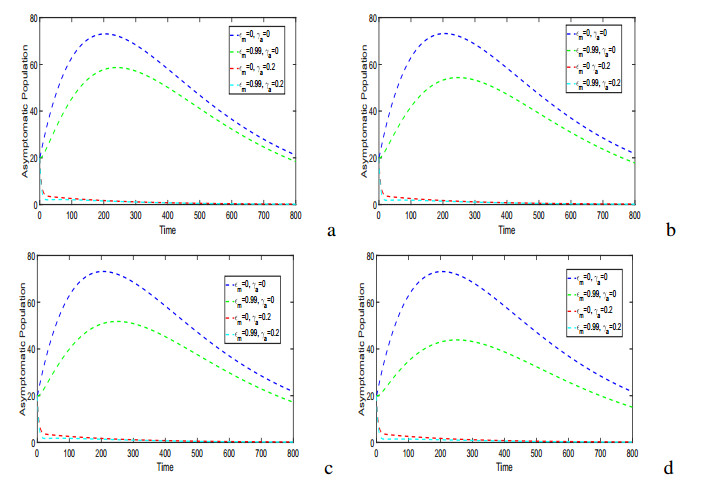
Figures(17) / Tables(5)

Akhil Kumar Srivastav, Pankaj Kumar Tiwari, Prashant K Srivastava, Mini Ghosh, Yun Kang. A mathematical model for the impacts of face mask, hospitalization and quarantine on the dynamics of COVID-19 in India: deterministic vs. stochastic[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 182-213. doi: 10.3934/mbe.2021010







 DownLoad:
DownLoad: