Citation: Xinran Zhou, Long Zhang, Tao Zheng, Hong-li Li, Zhidong Teng. Global stability for a class of HIV virus-to-cell dynamical model with Beddington-DeAngelis functional response and distributed time delay[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4527-4543. doi: 10.3934/mbe.2020250

| [1] | D. D. Richman, D. M. Margolis, M. Delaney, W. C. Greene, D. Hazuda, R. J. Pomerantz, The challenge of finding a cure for HIV infection, Science, 323 (2009), 1304-1307. |
| [2] | V. Leonenko, G. Bobashev, Analyzing influenza outbreaks in Russia using an age-structured dynamic transmission model, Epidemics, 29 (2019), 100-358. |
| [3] | Y. Cai, K. Wang, W. Wang, Global transmission dynamics of a Zika virus model, Appl. Math. Lett., 92 (2019), 190-195. |
| [4] | Y. Cai, Z. Ding, B. Yang, Z. Peng, W. Wang, Transmission dynamics of Zika virus with spatial structure-A case study in Rio de Janeiro, Brazil, Physica A: Stat. Mech. Appl., 514 (2019), 729-740. |
| [5] | E. Grigorieva, E. Khailov, Determination of the opimal controls for an Ebola epidemic model, Disc. Cont. Dynam. Syst. S., 11 (2018), 1071-1101. |
| [6] | J. Huang, S. Ruan, X. Wu, X. Zhou, Seasonal transmission dynamics of measles in China, Theor. Biosci., 137 (2018), 185-195. |
| [7] | T. Zhang, X. Zhao, Mathematical modeling for schistosomiasis with seasonal influence: A case study in Hubei, China, SIAM J. Appl. Dyn. Syst., 19 (2020), 1438-1471. |
| [8] | M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent virus, Science, 272 (1996), 74-79. |
| [9] | R. M. Anderson, R. M. May, The population dynamics of microparasites and their invertebrate hosts, Philos. Trans. R. Soc. Lond. Ser. B, 291 (1981), 451-524. |
| [10] | A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-I: dynamics in vivo, SIAM Rev., 41 (1999), 3-44. |
| [11] | X. Lai, X. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898-917. |
| [12] | A. L. Hill, D. I. Rosenbloom, M. A. Nowak, R. F. Siliciano, Insight into treatment of HIV infection from viral dynamics models, Immun. Rev., 285 (2018), 9-25. |
| [13] | P. Aavani, L. S. Allen, The role of CD4 T cells in immune system activation and viral reproduction in a simple model for HIV infection, Appl. Math. Model., 75 (2019), 210-222. |
| [14] | D. Olabode, L. Rong, X. Wang, Optimal control in HIV chemotherapy with termination viral load and latent reservoir, Math. Biosci. Eng., 16 (2018), 619-635. |
| [15] | X. Wang, L. Rong, HIV low viral load persistence under treatment: Insights from a model of cell-to-cell viral transmission, Appl. Math. Lett., 94 (2019), 44-51. |
| [16] | P. W. Nelson, A.S. Perelson, Mathematical analysis of delay differential equation models of HIV-1 infection, Math. Biosci., 179 (2002), 73-94. |
| [17] | R. Xu, Global stability of an HIV-1 infection model with saturation infection and intracellular delay, J. Math. Anal. Appl., 375 (2011), 75-81. |
| [18] | J. Lin, R. Xu, X, Tian, Threshold dynamics of an HIV-1 virus model with both virus-to-cell and cell-to-cell transmissions, intracellular delay, and humoral immunity, Appl. Math. Comput., 315 (2017), 516-530. |
| [19] | G. Huang, W. Ma, Y. Takeuchi, Global properties for virus dynamics model with BeddingtonDeAngelis functional response, Appl. Math. Lett., 22 (2009), 1690-1693. |
| [20] | G. Huang, W. Ma, Y. Takeuchi, Global analysis for delay virus dynamics model with BeddingtonDeAngelis functional response, Appl. Math. Lett., 24 (2011), 1199-1203. |
| [21] | Y. Nakata, Global dynamics of a viral infection model with a latent period and beddingtonDeAngelis response, Nonlinear Anal. TMA, 74 (2011), 2929-2940. |
| [22] | H. Xiang, L. Feng, H. Huo, Stability of the virus dynamics model with Beddington-DeAngelis functional response and delays, Appl. Math. Model., 37 (2013), 5414-5423. |
| [23] | R. Xu, Global dynamics of an HIV-1 infection model with distributed intracellular delays, Comput. Math. Appl., 61 (2011), 2799-2805. |
| [24] | J. Wang, M. Guo, X. Liu, Z. Zhao, Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149-161. |
| [25] | Y. Nakata, Global dynamics of a cell mediated immunity in viral infection models with distributed delays, J. Math. Anal. Appl., 375 (2011), 14-27. |
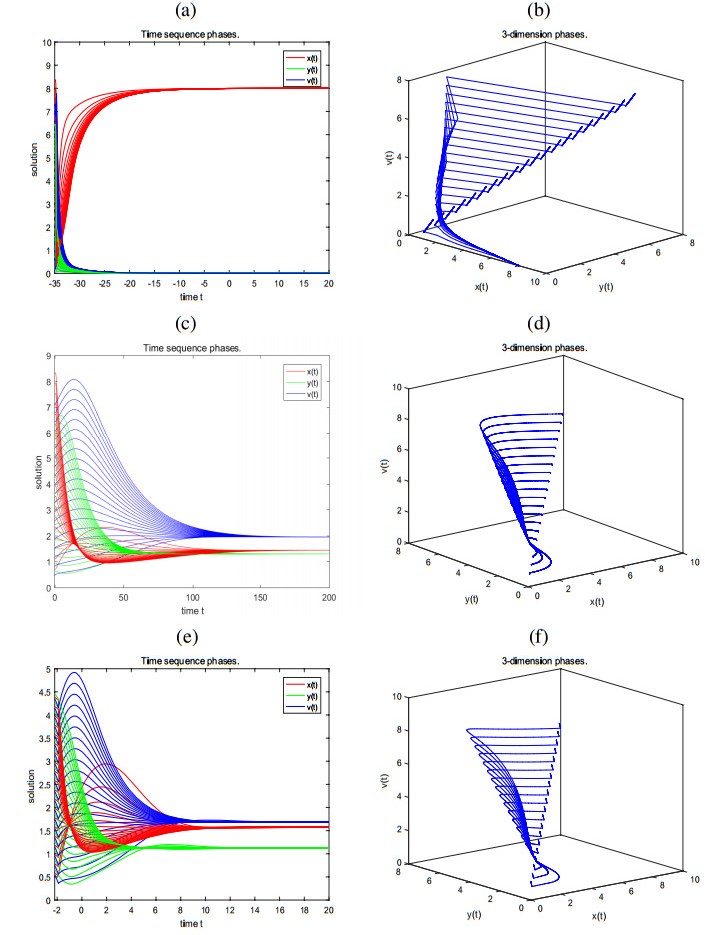
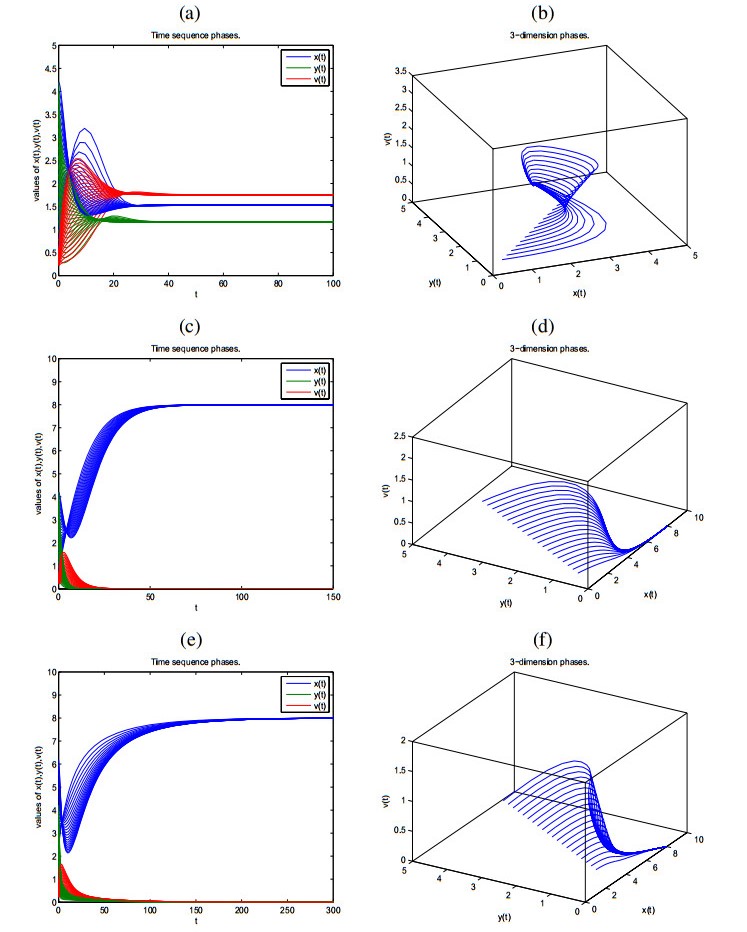
Figures(2) / Tables(2)

Xinran Zhou, Long Zhang, Tao Zheng, Hong-li Li, Zhidong Teng. Global stability for a class of HIV virus-to-cell dynamical model with Beddington-DeAngelis functional response and distributed time delay[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4527-4543. doi: 10.3934/mbe.2020250







 DownLoad:
DownLoad: