Citation: Ran Zhang, Shengqiang Liu. Global dynamics of an age-structured within-host viral infection model with cell-to-cell transmission and general humoral immunity response[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1450-1478. doi: 10.3934/mbe.2020075

| [1] | M. A. Nowak, S. Bonhoeffer, A. M. Hill, R. Boehme, H. C. Thomas, H. McDade, Viral dynamics in hepatitis B virus infection, Proc. Natl. Acad. Sci. USA, 93 (1996), 4398-4402. |
| [2] | P. Nelson, M. Gilchrist, D. Coombs, J. M. Hyman, A. S. perelson, An age-structured model of HIV infection that allow for variations in the production rate of viral particles and the death rate of productively infected cells, Math. Biosci. Eng., 1 (2004), 267-288. |
| [3] | L. Rong, Z. Feng, A. S. Perelson, Mathematical analysis of age-structured HIV-1 dynamics with combination antiretroviral therapy, SIAM J. Appl. Math., 67 (2007), 731-756. |
| [4] | G. Huang, X. Liu, Y. Takeuchi, Lyapunov functions and global stability for age-structured HIV infection model, SIAM J. Appl. Math., 72 (2012), 25-38. |
| [5] | L. Zou, S. Ruan, W. Zhang, An age-structured model for the transmission dynamics of hepatitis B, SIAM J. Appl. Math., 70 (2010), 3121-3139. |
| [6] | J. Wang, R. Zhang, T. Kuniya, Global dynamics for a class of age-infection HIV models with nonlinear infection rate, J. Math. Anal. Appl., 432 (2015), 289-313. |
| [7] | M. Shen, Y. Xiao, L. Rong, Global stability of an infection-age structured HIV-1 model linking within-host and between-host dynamics, Math. Biosci., 263 (2015), 37-50. |
| [8] | Y. Wang, K. Liu, Y. Lou, An age-structured within-host HIV model with T-cell competition, Nonlinear Anal. Real World Appl., 38 (2017), 1-20. |
| [9] | C.-Y. Cheng, Y. Dong, Y. Takeuchi, An age-structured virus model with two routes of infection in heterogeneous environments, Nonlinear Anal. Real World Appl., 39 (2018), 464-491. |
| [10] | Z. Liu, Y. Rong, Zero-Hopf bifurcation for an infection-age structured epidemic model with a nonlinear incidence rate, Sci. China Math., 60 (2017), 1371-1398. |
| [11] | J. Wang, X. Dong, Analysis of an HIV infection model incorporating latency age and infection age, Math. Biosci. Eng., 15 (2018), 569-594. |
| [12] | J. Pang, J. Chen, Z. Liu, P. Bi, S. Ruan, Local and global stabilities of a viral dynamics model with infection-age and immune response, J. Dyn. Diff. Equat., 31 (2019), 793-813. |
| [13] | K. Hattaf, Y. Yang, Global dynamics of an age-structured viral infection model with general incidence function and absorption, Int. J. Biomath., 11 (2018), 1850065. |
| [14] | W. Hübner, G. McNerney, P. Chen, B. M. Dale, R. E. Gordon, F. Y. Chuang, et al., Quantitative 3D video microscopy of HIV transfer across T cell virological synapses, Science, 323 (2009), 1743-1747. |
| [15] | A. Imle, P. Kumberger, N. D. Schnellbächer, J. Fehr, P. Carrillo-Bustamante, J. Ales, et al., Experimental and computational analyses reveal that environmental restrictions shape HIV-1 spread in 3D cultures, Nat. Commun., 10 (2019), 2144. |
| [16] | N. Barretto, B. Sainz, S. Hussain, S. L. Uprichard, Determining the involvement and therapeutic implications of host cellular factors in hepatitis C virus cell-to-cell spread, J. Virol., 88 (2014), 5050-5061. |
| [17] | F. Merwaiss, C. Czibener, D. E. Alvarez, Cell-to-cell transmission is the main mechanism supporting bovine viral diarrhea virus spread in cell culture, J. Virol., 93 (2019), e01776. |
| [18] | G. L. Smith, B. J. Murphy, M. Law, Vaccinia virus motility, Annu. Rev. Microbiol., 57 (2003), 323-342. |
| [19] | X. Lai, X. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898-917. |
| [20] | Y. Yang, L. Zou, S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183-191. |
| [21] | J. Wang, J. Lang, X. Zou, Analysis of an age structured HIV infection model with virus-to-cell infection and cell-to-cell transmission, Nonlinear Anal. Real World Appl., 34 (2017), 75-96. |
| [22] | X. Zhang, Z. Liu, Bifurcation analysis of an age structured HIV infection model with both virusto-cell and cell-to-cell transmissions, Int. J. Bifurcation Chaos, 28 (2018), 1850109. |
| [23] | A. Murase, T. Sasaki, T. Kajiwara, Stability analysis of pathogen-immune interaction dynamics, J. Math. Biol., 51 (2005), 247-267. |
| [24] | S. Wang, D. Zou, Global stability of in-host viral models with humoral immunity and intracellular delays, Appl. Math. Model, 51 (2012), 1313-1322. |
| [25] | T. Wang, Z. Hu, F. Liao, Stability and Hopf bifurcation for a virus infection model with delayed humoral immunity response, J. Math. Appl. Anal., 411 (2014), 63-74. |
| [26] | T. Kajiwara, T. Sasaki, Y. Otani, Global stability of an age-structured model for pathogen-immune interaction, J. Appl. Math. Comput., 59 (2019), 631-660. |
| [27] | M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. |
| [28] | Y. Wang, Y. Zhou, F. Brauer, J. M. Heffernan, Viral dynamics model with CTL immune response incorporating antiretroviral therapy, J. Math. Biol., 67 (2013), 901-934. |
| [29] | C. Browne, Immune response in virus model structured by cell infection-age, Math. Biosci. Eng., 13 (2016), 887-909. |
| [30] | X. Duan, S. Yuan, Global dynamics of an age-structured virus model with saturation effects, Math. Methods Appl. Sci., 40 (2017), 1851-1864. |
| [31] | X. Wang, S. Liu, A class of delayed viral models with saturation infection rate and immune response, Math. Methods Appl. Sci., 36 (2013), 125-142. |
| [32] | H. Shu, L. Wang, J. Watmough, Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL immune responses, SIAM J. Appl. Math., 73 (2013), 1280-1302. |
| [33] | X. Tian, R. Xu, J. Lin, Mathematical analysis of an age-structured HIV-1 infection model with CTL immune response, Math. Biosci. Eng., 16 (2019), 7850-7882. |
| [34] | K. Hattaf, Spatiotemporal dynamics of a generalized viral infection model with distributed delays and CTL immune response, Computation, 7 (2019), 21. |
| [35] | X. Tian, R. Xu, Global stability of a delayed HIV-1 infection model with absorption and CTL immune response, IMA J. Appl. Math., 79 (2014), 347-359. |
| [36] | P. Magal, S. Ruan, Theory and Applications of Abstract Semilinear Cauchy Problems, Applied Mathematical Sciences Vol. 201, Springer, Cham, 2018. |
| [37] | P. Magal, Compact attractors for time-periodic age structured population models, Elect. J. Differ. Eqs., 65 (2001), 65. |
| [38] | P. Magal, C. C. McCluskey, G. F. Webb, Lyapunov functional and global asymptotic stability for an infection-age model, Appl. Anal., 89 (2010), 1109-1140. |
| [39] | P. Magal, H. Thieme, Eventual compactness for a semiflow generated by an age-structured models, Commun. Pure Appl. Anal., 3 (2004), 695-727. |
| [40] | H. L. Smith, H. R. Thieme, Dynamical Systems and Population Persistence, Graduate Studies in Mathematics Vol. 118, American Mathematical Society, Providence, RI, 2011. |
| [41] | C. C. McCluskey, Global stability for an SEI epidemiological model with continuous age-structure in the exposed and infectious classes, Math. Biosci. Eng., 9 (2012), 819-841. |
| [42] | C. Jiang, K. Wang, L. Song, Global dynamics of a delay virus model with recruitment and saturation effects of immune responses, Math. Biosci. Eng., 14 (2017), 1233-1246. |
| [43] | H. Dahari, A. Lo, R. M. Ribeiro, A. S. Perelson, Modeling hepatitis C virus dynamics: liver regeneration and critical drug efficacy, J. Theor. Biol., 247 (2007), 371-381. |
| [44] | Y. Wang, Y. Zhou, J. Wu, J. Heffernan, Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104-112. |
| [45] | J. Reyes-Silveyra, A. R. Mikler, Modeling immune response and its effect on infectious disease outbreak dynamics, Theor. Biol. Med. Model., 13 (2016), 10. |
| [46] | R. Qesmi, S. ElSaadany, J. M. Heffernan, J. Wu, A hepatitis B and C virus model with age since infection that exhibits backward bifurcation, SIAM J. Appl. Math., 71 (2011), 1509-1530. |
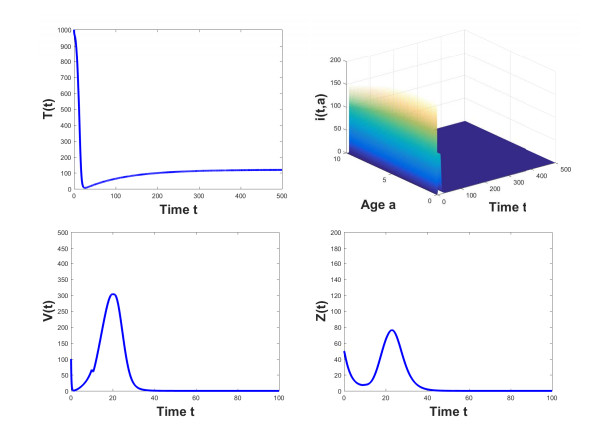
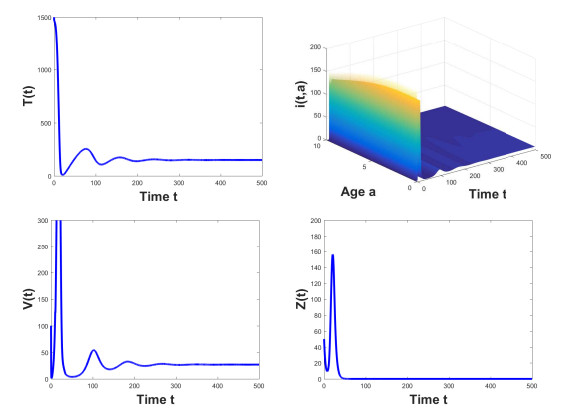
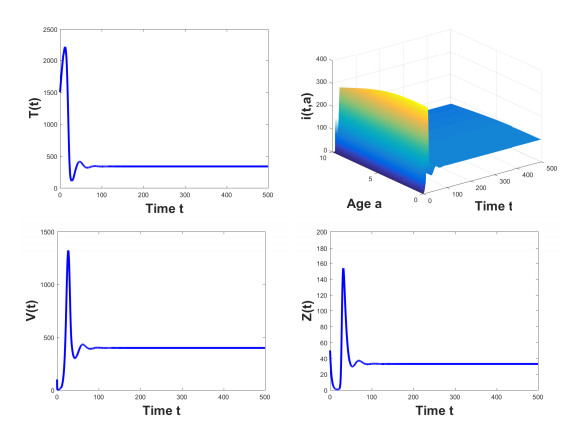
Figures(3) / Tables(2)

Ran Zhang, Shengqiang Liu. Global dynamics of an age-structured within-host viral infection model with cell-to-cell transmission and general humoral immunity response[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1450-1478. doi: 10.3934/mbe.2020075







 DownLoad:
DownLoad: