Citation: Antonella Lupica, Piero Manfredi, Vitaly Volpert, Annunziata Palumbo, Alberto d'Onofrio. Spatio-temporal games of voluntary vaccination in the absence of the infection: the interplay of local versus non-local information about vaccine adverse events[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1090-1131. doi: 10.3934/mbe.2020058

| [1] | N. T. J. Bailey, The mathematical theory of infectious diseases and its applications (2nd edition), 2nd edition, Charles Griffin & Company Ltd, 1975. |
| [2] | V. Capasso and V. Capasso, Mathematical structures of epidemic systems, Springer, 1993. |
| [3] | C. Metcalf, E. Jessica, W. J. Edmunds, et al., Six challenges in modelling for public health policy, Epidemics, 10 (2015), 93-96. |
| [4] | P. Manfredi and A. d'Onofrio, Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases, Springer, 2013. |
| [5] | Z. Wang, C. T. Bauch, S. Bhattacharyya, et al., Statistical physics of vaccination, Phys. Rep., 664 (2016), 1-113. |
| [6] | V. Capasso and G. Serio, A generalization of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43-61. |
| [7] | R. Casiday, T. Cresswell, D. Wilson, et al., A survey of UK parental attitudes to the MMR vaccine and trust in medical authority, Vaccine, 24 (2006), 177-184. |
| [8] | E. Dubé, C. Laberge, M. Guay, et al., Vaccine hesitancy: an overview, Human Vaccin. Immunother., 9 (2013), 1763-1773. |
| [9] |
V. A. A. Jansen, N. Stollenwerk, H. J. Jensen, et al., Measles outbreaks in a population with declining vaccine uptake, Science, 301 (2003), 804-804. doi: 10.1126/science.1086726

|
| [10] | S. B. Omer, D. A. Salmon, W. A. Orenstein, et al., Vaccine refusal, mandatory immunization, and the risks of vaccine-preventable diseases, N. Engl. J. Med., 360 (2009), 1981-1988. |
| [11] | R. Löfstedt, Risk management in post-trust societies, Palgrave-McMillan, 2005. |
| [12] | C. T. Bauch, Imitation dynamics predict vaccinating behaviour, Proc. R. Soc. Lond. B Biol. Sci., 272 (2005), 1669-1675. |
| [13] | A. d'Onofrio, P. Manfredi and P. Poletti, The impact of vaccine side effects on the natural history of immunization programmes: an imitation-game approach, J. Theor. Biol., 273 (2011), 63-71. |
| [14] | A. d'Onofrio, P. Manfredi and P. Poletti, The interplay of public intervention and private choices in determining the outcome of vaccination programmes, PLoS ONE, 7 (2012), e45653. |
| [15] | B. Buonomo, G. Carbone and A. d'Onofrio, Effect of seasonality on the dynamics of an imitation- based vaccination model with public health intervention, Math. Biosci. Eng., 15 (2018), 299-321. |
| [16] | D. G. Kendall, Mathematical models of the spread of infection, Math. Comput. Sci. Biol. Med. (1965), 213-225. |
| [17] | N. T. J. Bailey, The simulation of stochastic epidemics in two dimensions, Proc. Fifth Berkeley Symp. Math. Statist. Prob., 4 (1967), 237-257. |
| [18] | D. G. Aronson, The asymptotic speed of propagation of a simple epidemic, Nonlinear Diffusion, 14 (1977), 1-23. |
| [19] | V. Capasso, Global solution for a diffusive nonlinear deterministic epidemic model, SIAM J. Appl. Math., 35 (1978), 274-284. |
| [20] | F. Hoppensteadt, Mathematical Theories of Populations: Deomgraphics, Genetics, and Epidemics, Siam, 1975. |
| [21] | H. Malchow, S. V. Petrovskii and E. Venturino, Spatiotemporal patterns in ecology and epidemiology: theory, models, and simulation, Chapman and Hall/CRC, 2007. |
| [22] | J. D. Murray, Mathematical biology II. Spatial models and biomedical applications, SpringerVerlag New York Incorporated New York, 2001. |
| [23] | S. Ruan and J. Wu, Modeling spatial spread of communicable diseases involving animal hosts, Spatial Ecol., (2009), 293-316. |
| [24] | J. V. Noble, Geographic and temporal development of plagues, Nature, 250 (1974), 726. |
| [25] | A. Ducrot and T. Giletti, Convergence to a pulsating travelling wave for an epidemic reactiondiffusion system with non-diffusive susceptible population, J. Math. Biol., 69 (2014), 533-552. |
| [26] | P. Magal, G. F. Webb and Y. Wu, On the Basic Reproduction Number of Reaction-Diffusion Epidemic Models, SIAM J. Appl. Math., 79 (2019), 284-304. |
| [27] | P. Magal, G. F. Webb and Y. Wu, Spatial spread of epidemic diseases in geographical settings: seasonal influenza epidemics in Puerto Rico, arXiv preprint arXiv:1801.01856 (2018). |
| [28] | W. E. Fitzgibbon, J. J. Morgan, G. F. Webb, et al., A vector-host epidemic model with spatial structure and age of infection, Nonlinear Anal-Real., 41 (2018), 692-705. |
| [29] | L. Zhao, Z. C. Wang and S. Ruan, Traveling wave solutions in a two-group SIR epidemic model with constant recruitment, J. Math. Biol., 77 (2018), 1871-1915. |
| [30] | S. Ruan, Spatial-temporal dynamics in nonlocal epidemiological models, Math. Life Sci. Med., (2007), 97-122. |
| [31] | B. Buonomo, P. Manfredi and A. d'Onofrio, Optimal time-profiles of public health intervention to shape voluntary vaccination for childhood diseases, J. Math. Biol., 78 (2019), 1089-1113. |
| [32] | R. Peres, E. Muller and V. Mahajan, Innovation diffusion and new product growth models: A critical review and research directions, Int. J. Res. Market., 27 (2010), 91-106. |
| [33] | V. Capasso and M. Zonno, Mathematical Models for the Diffusion of Innovations, Proc. Fourth Eur. Conference Math. Industry, (1991), 225-233. |
| [34] | V. Mahajan and R. A. Peterson, Models for innovation diffusion, Sage Publications Inc, 1985. |
| [35] | V. Capasso, A. Di Liddo and L. Maddalena, A nonlinear model for the geographical spread of innovations, Dyn. Syst Appl., 3 (1994), 207-220. |
| [36] | F. M. Bass, A new product growth for model consumer durables, Manag. Sci., 15 (1969), 215-227. |
| [37] | S. Funk, E. Gilad, C. Watkins, et al., The spread of awareness and its impact on epidemic outbreaks, Proc. Natl. Acad. Sci., 106 (2009), 6872-6877. |
| [38] | E. Frey, Evolutionary game theory: Theoretical concepts and applications to microbial communities, Physica A, 389 (2010), 4265-4298. |
| [39] | R. A. Fisher, The wave of advance of advantageous genes, Annals Eugenics, 7 (1937), 355-369. |
| [40] | V. Volpert, Elliptic Partial Differential Equations: Volume 2: Reaction-Diffusion Equations, 104, Springer, 2014. |
| [41] | A. N. Kolmogorov, I. N. Petrovsky and N. S. Piskunov, Étude de l'équation de la diffusion avec croissance de la quantité de matière et son application à un problème biologique, Bull. Univ. Moskow, Ser. Internat., Sec. A, 1 (1937), 1-25. |
| [42] | A. D. Polyanin and V. F. Zaitsev, Handbook of ordinary differential equations: exact solutions, methods, and problems, Chapman and Hall/CRC, 2017. |
| [43] | J. D. Murray, Mathematical biology I: an introduction, Springer New York, 2002. |
| [44] | A. I. Volpert, V. A. Volpert and V. A. Volpert, Traveling wave solutions of parabolic systems, 140 American Mathematical Soc., 1994. |
| [45] | S. Vakulenko and V. Volpert, Generalized travelling waves for perturbed monotone reactiondiffusion systems, Nonlinear Analysis TMA, 6 (2001), 757-776. |
| [46] | A. d'Onofrio, P. Manfredi and E. Salinelli, Vaccinating behaviour and the dynamics of vaccine preventable infections, Modeling the Interplay Between Human Behavior and the Spread of Infectious Diseases (2013), 267-287. |
| [47] | M. Mincheva and M. R. Roussel, Turing-Hopf instability in biochemical reaction networks arising from pairs of subnetworks, Math. Biosci., 240 (2012), 1-11. |
| [48] | B. Ermentrout and M. Lewis, Pattern formation in systems with one spatially distributed species, Bull. Math. Biol., 59 (1997), 533-549. |
| [49] | A. M. Turing, The chemical basis of morphogenesis, Phil. Trans. R. Soc. Lond. B, 237 (1952), 37-72. |
| [50] | N. Stollenwerk and V. Jansen, Population Biology and Criticality: From critical birth-death processes to self-organized criticality in mutation pathogen systems, World Scientific, 2011. |
| [51] | N. Stollenwerk, S. van Noort, J. Martins, et al., A spatially stochastic epidemic model with partial immunization shows in mean field approximation the reinfection threshold, J. Biol. Dyn., 4 (2010), 634-649. |
| [52] | A. M. Albano, N. B. Abraham, D. E. Chyba, et al., Bifurcations, propagating solutions, and phase transitions in a nonlinear chemical reaction with diffusion, Am. J. Phys., 52 (1984), 161-167. |
| [53] | J. P. Eckmann and T. Gallay, Front solutions for the Ginzburg-Landau equation, Commun. Math. Phys., 152 (1993), 221-248. |
| [54] | B. J. Matkowsky and V. A. Volpert, Stability of plane wave solutions of complex Ginzburg-Landau equations, Quart. Appl. Math., 51 (1993), 265-281. |
| [55] | L. E. Reichl, A modern course in statistical physics, Wiley-VCH, 2016. |
| [56] | M. C. Gonzalez, C. A. Hidalgo and A. L. Barabasi, Understanding individual human mobility patterns, Nature, 453 (2008), 779. |
| [57] | D. Balcan and A. Vespignani, Phase transitions in contagion processes mediated by recurrent mobility patterns, Nature Phys., 7 (2011), 581. |
| [58] | U. Skwara, J. Martins, P. Ghaffari, et al., Fractional calculus and superdiffusion in epidemiology: shift of critical thresholds, Proceedings of the 12th International Conference on Computational and Mathematical Methods in Science and Engineering, La Manga (2012). |
| [59] | J. P. Boto and N. Stollenwerk, Fractional calculus and Levy flights: modelling spatial epidemic spreading, Computational and Mathematical Methods in Science and Engineering, (2009). |
| [60] | U. Skwara, J. Martins, P. Ghaffari, et al., Applications of fractional calculus to epidemiological models, AIP Conf. Proc., 1479 (2012), 1339-1342. |
| [61] | U. Skwara, L. Mateus, R. Filipe, et al., Superdiffusion and epidemiological spreading, Ecol. Complex., 36 (2018), 168-183. |
| [62] | D. Brockmann and L. Hufnagel, Front propagation in reaction-superdiffusion dynamics: Taming Lévy flights with fluctuations, Phys. Rev. Lett., 98 (2009), 178301. |
| [63] | D. R. Sinclair, J. J. Grefenstette, M. G. Krauland, et al., Forecasted Size of Measles Outbreaks Associated With Vaccination Exemptions for Schoolchildren, JAMA Network Open, 2 (2019), e199768-e199768. |
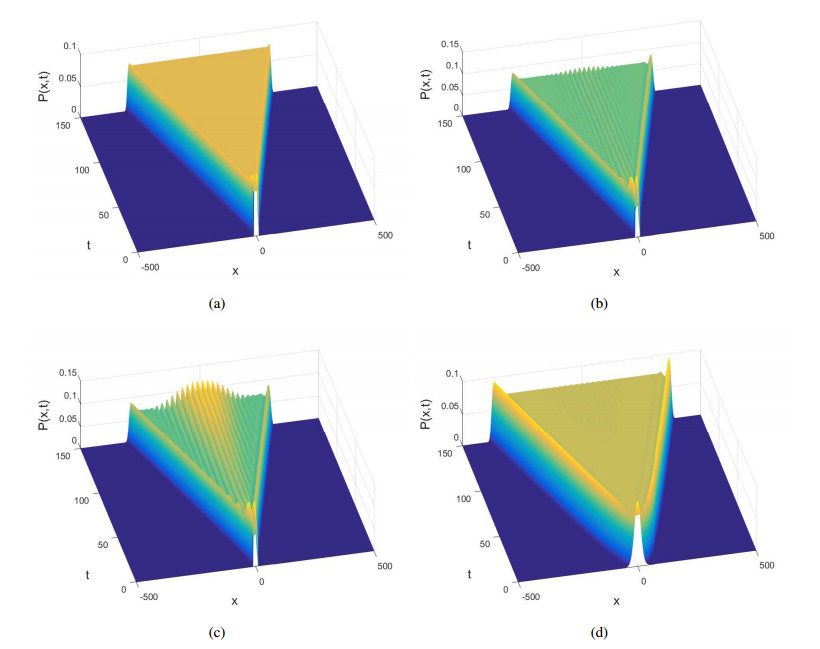
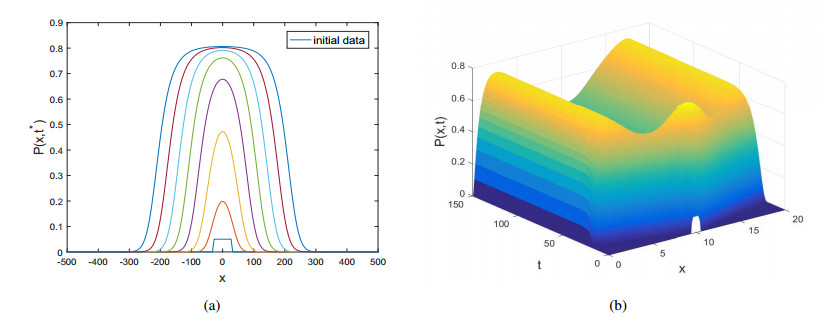
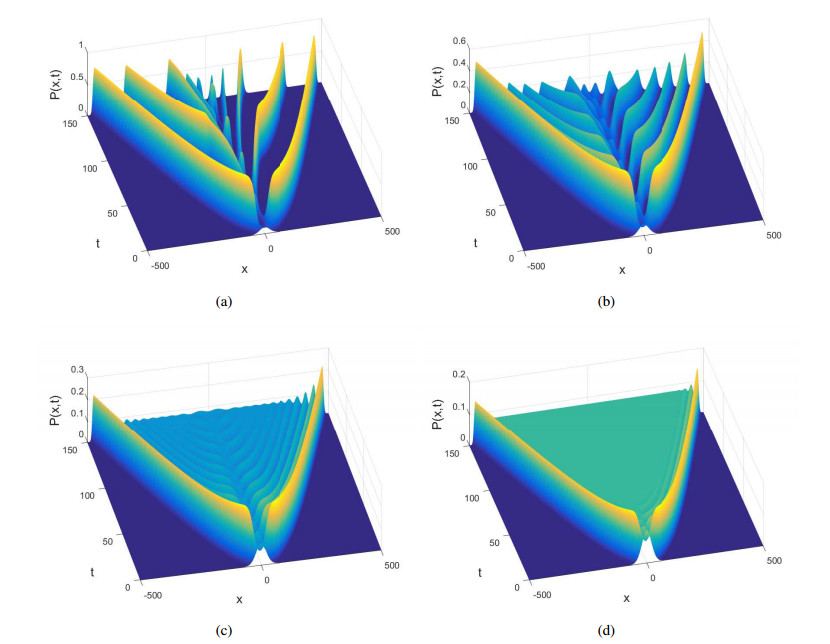
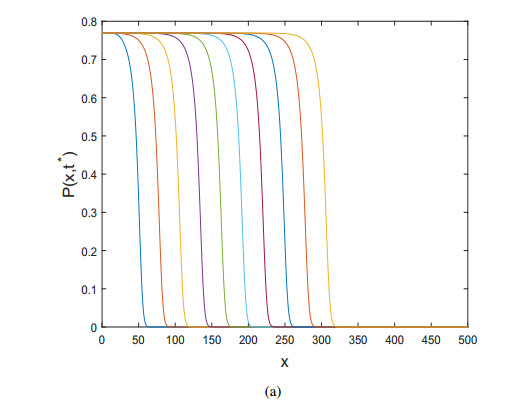
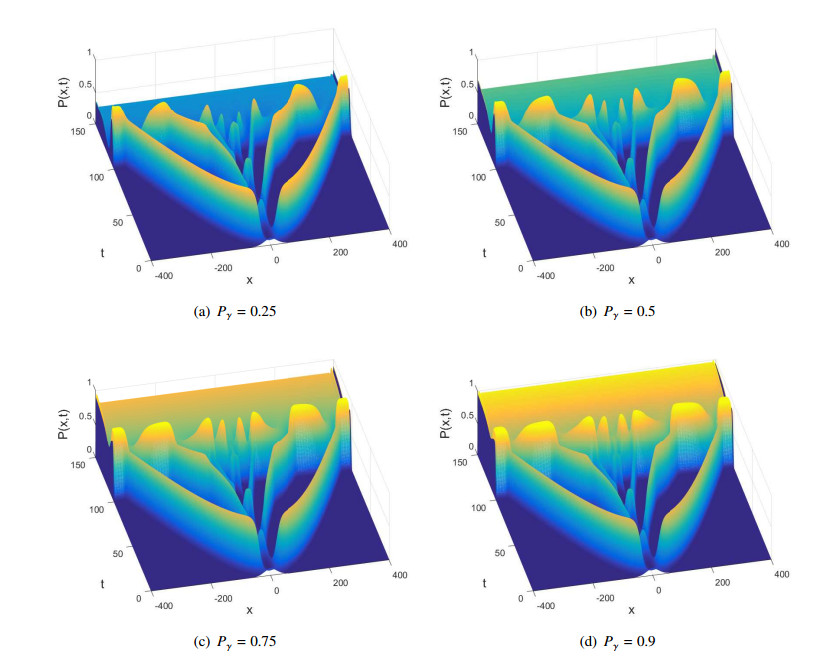
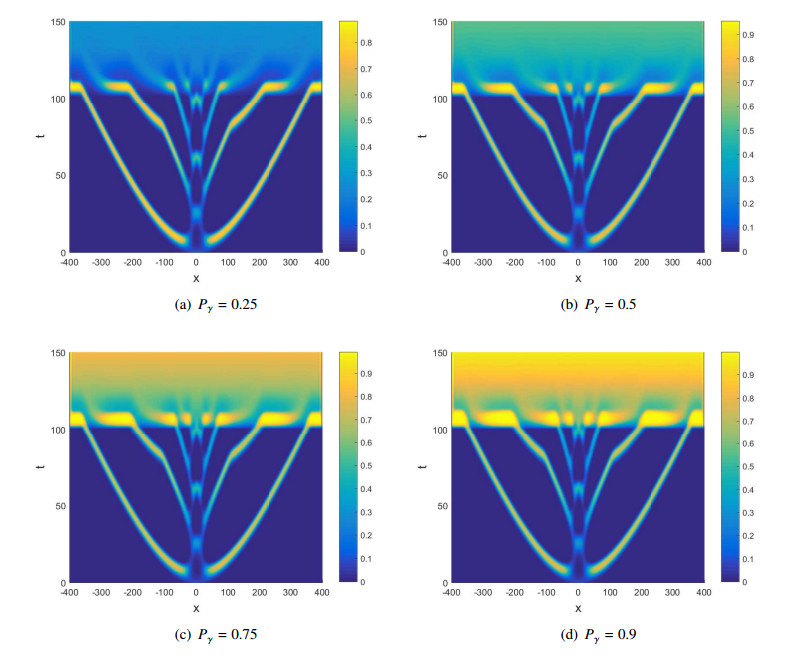
Figures(10)

Antonella Lupica, Piero Manfredi, Vitaly Volpert, Annunziata Palumbo, Alberto d'Onofrio. Spatio-temporal games of voluntary vaccination in the absence of the infection: the interplay of local versus non-local information about vaccine adverse events[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1090-1131. doi: 10.3934/mbe.2020058







 DownLoad:
DownLoad: