Citation: Zhong-Kai Guo, Hai-Feng Huo, Hong Xiang. Global dynamics of an age-structured malaria model with prevention[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1625-1653. doi: 10.3934/mbe.2019078

| [1] | World Health Organization,World malaria report, 2016. Available from: http://www.who.int/ malaria/publications/world-malaria-report-2016/en/. |
| [2] | CDC. malaria, malaria worldwide, 2018. Available from: https://www.cdc.gov/malaria/ about/biology/index.html. |
| [3] | R. Ross, The Prevention of Malaria, 2nd edition, Murray, London, 1911. |
| [4] | G. Macdonald, The epidemiology and control of malaria, Oxford University press, London, 1957. |
| [5] | S. A. Gourley, R. Liu and J. Wu, Some vector borne diseases with structured host populations: Extinction and spatial spread, SIAM J. Appl. Math., 67 (2007), 408–433. |
| [6] | Fatmawati and H. Tasman, An optimal control strategy to reduce the spread of malaria resistance, Math. Biosci., 262 (2015), 73–79. |
| [7] | S. Ruan, D. Xiao and J. C. Beier, On the delayed Ross-Macdonald model for malaria transmission, B. Math. Biol., 70 (2008), 1098–1114. |
| [8] | L. M. Cai, X. Z. Li, B. Fang, et al., Global properties of vector–host disease models with time delays, J. Math. Biol., 74 (2017), 1397–1423. |
| [9] | J. Xu and Y. Zhou, Hopf bifurcation and its stability for a vector-borne disease model with delay and reinfection, Appl. Math. Model., 40 (2016), 1685–1702. |
| [10] | Y. Lou and X. Q. Zhao, A climate-based malaria transmission model with structured vector population, SIAM J. Appl. Math., 70 (2010), 2023–2044. |
| [11] | Y. X. Dang, Z. P. Qiu, X. Z. Li, et al., Global dynamics of a vector-host epidemic model with age of infection, Math. Biosci. Eng., 14 (2017), 1159–1186. |
| [12] | D. Gao and S. Ruan, A multipatch malaria model with logistic growth populations, SIAM J. Appl. Math., 72 (2012), 819–841. |
| [13] | J. Li, Y. Zhao and S. Li, Fast and slow dynamics of malaria model with relapse, Math. Biosci., 246 (2013), 94–104. |
| [14] | B. Cao, H. F. Huo and H. Xiang, Global stability of an age-structure epidemic model with imperfect vaccination and relapse, Physica A, 486 (2017), 638–655. |
| [15] | X. Duan, S. Yuan and X. Li, Global stability of an SVIR model with age of vaccination, Appl. Math. Comput., 226 (2014), 528–540. |
| [16] | H. Inaba and H. Sekine, A mathematical model for chagas disease with infection-age-dependent infectivity, Math. Biosci., 190 (2004), 39–69. |
| [17] | L.Wang, Z. Liu and X. Zhang, Global dynamics for an age-structured epidemic model with media impact and incomplete vaccination, Nonlinear Anal. RWA, 32 (2016), 136–158. |
| [18] | H. Chen, W. Wang, R. Fu, et al., Global analysis of a mathematical model on malaria with competitive strains and immune responses, Appl. Math. Comput., 259 (2015), 132–152. |
| [19] | H. Xiang, Y. L. Tang and H. F. Huo, A viral model with intracellular delay and humoral immunity, B. Malays. Math. Sci. So., 40 (2017), 1011–1023. |
| [20] | H. F. Huo, F. F. Cui and H. Xiang, Dynamics of an SAITS alcoholism model on unweighted and weighted networks, Physica A, 496 (2018), 249–262. |
| [21] | H. Xiang, Y. Y. Wang and H. F. Huo, Analysis of the binge drinking models with demographics and nonlinear infectivity on networks, J. Appl. Anal. Comput., 8 (2018), 1535–1554. |
| [22] | Y. Cai, J. Jiao, Z. Gui, et al., Environmental variability in a stochastic epidemic model, Appl. Math. Comput., 329 (2018), 210–226. |
| [23] | X. B. Zhang, Q. H. Shi, S. H. Ma, et al., Dynamic behavior of a stochastic SIQS epidemic model with Lévy jumps, Nonlinear Dynam., 93 (2018), 1481–1493. |
| [24] | Z. Du and Z. Feng, Existence and asymptotic behaviors of traveling waves of a modified vectordisease model, Commun. pur. appl. anal., 17 (2018), 1899–1920. |
| [25] | Malaria vaccine. Available from: https://en.wikipedia.org/wiki/Malaria_vaccine. |
| [26] | G. Webb, Theory of nonlinear age-dependent population dynamics, marcel dekker, New York, 1985. |
| [27] | M. Iannelli, Mathematical theory of age-structured population dynamics, Giadini Editori e Stampatori, Pisa, 1994. |
| [28] | H. L. Smith and H. R. Thieme, Dynamical systems and population persistence, American Mathematical Society, Providence, 2011. |
| [29] | P. Magal and X. Q. Zhao, Global attractors and steady states for uniformly persistent dynamical systems, SIAM J. Appl. Math., 37 (2005), 251–275. |
| [30] | E. M. C. D'Agata, P. Magal, S. Ruan, et al., Asymptotic behavior in nosocomial epidemic models with antibiotic resistance, Differ. Integral Equ., 19 (2006), 573–600. |
| [31] | C. J. Browne and S. S. Pilyugin, Global analysis of age-structured within-host virus model, Discrete Cont. Dyn. S. B, 18 (2013), 1999–2017. |
| [32] | H. M. Wei, X. Z. Li and M. Martcheva, An epidemic model of a vector-borne disease with direct transmission and time delay, J. Math. Anal. Appl., 342 (2008), 895–908. |
| [33] | J. Wang, J. Lang and X. Zou, Analysis of an age structured hiv infection model with virus-to-cell infection and cell-to-cell transmission, Nonlinear Anal. RWA, 34 (2017), 75–96. |
| [34] | J. K. Hale and P. Waltman, Persistence in infinite-dimensional systems, SIAM J. Appl. Math., 20 (1989), 388–395. |
| [35] | Z. K. Guo, H. F. Huo, X. Hong, Bifurcation analysis of an age-structured alcoholism model, J. Biol. Dynam., 12 (2018), 1009–1033. |
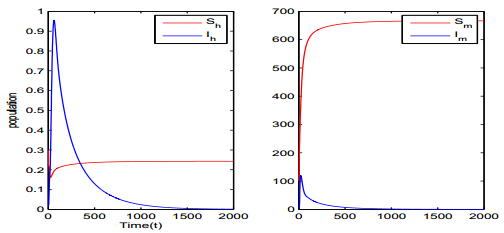
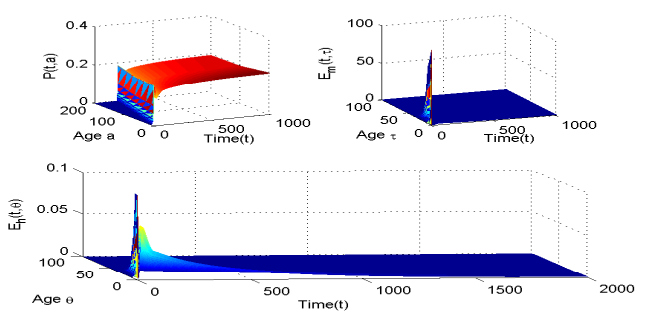
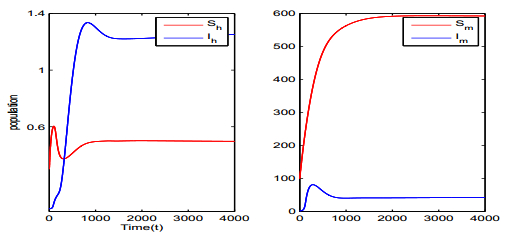
Figures(5) / Tables(1)

Zhong-Kai Guo, Hai-Feng Huo, Hong Xiang. Global dynamics of an age-structured malaria model with prevention[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1625-1653. doi: 10.3934/mbe.2019078







 DownLoad:
DownLoad: