Citation: Md Nazmul Hassan, Kelsey Thompson, Gregory Mayer, Angela Peace. Effect of Excess Food Nutrient on Producer-Grazer Model under Stoichiometric and Toxicological Constraints[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 150-167. doi: 10.3934/mbe.2019008

| [1] | T. Andersen, Pelagic nutrient cycles: herbivores as sources and sinks, 129 (2013), Springer Science & Business Media. |
| [2] | K.E. Biesinger, L.E. Anderson and J.G. Eaton, Chronic effects of inorganic and organic mercury ondaphnia magna: Toxicity, accumulation, and loss, Arc. Environment. Contamin. Toxico., 11 (1982), 769–774. |
| [3] | M. Boersma and J.J. Elser, Too much of a good thing: on stoichiometrically balanced diets and maximal growth, Ecology, 87 (2006), 1325–1330. |
| [4] | A.J. Cease, J.J. Elser, C.F. Ford, S. Hao, L. Kang and J.F. Harrison, Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content, Science, 335 (2012), 467–469. |
| [5] | M. Danger and F. Maunoury-Danger, Ecological stoichiometry. In: Encyclopedia of Aquatic Ecotoxicology, Springer, (2013), 317–326. |
| [6] | J.J. Elser, J. Watts, J.J. Schampell and J. Farmer, Early Cambrian food webs on a trophic knifeedge? a hypothesis and preliminary data from a modern stromatolite-based ecosystem, Ecol. Lett., 9 (2006), 295–303. |
| [7] | J.J. Elser, M. Kyle, J. Learned, M.L. McCrackin, A. Peace and L. Steger, Life on the stoichiometric knife-edge: effects of high and low food c: P ratio on growth, feeding, and respiration in three daphnia species, Inland Water., 6 (2006), 136–146. |
| [8] | L.K. Hansen, P.C. Fros, J.H. Larson and C.D. Metcalfe, Poor elemental food quality reduces the toxicity of fluoxetine on daphnia magna, Aquat. Toxicol., 86 (2008), 99–103. |
| [9] | W.R. Hill and I.L. Larsen, Growth dilution of metals in microalgal biofilms, Environment. Sci. Technol., 39 (2005), 1513–1518. |
| [10] | Q. Huang, H. Wang and M. Lewis, Development of a toxin-mediated predator-prey model applicable to aquatic environments in the athabasca oil sands region, Osrin Rep. Tech. Rep., 59 (2014), 55. http://hdl. handle. net/10402/era. 40140. |
| [11] | O. Ieromina, W.J. Peijnenburg, G. de Snoo, J. M¨uller, T.P. Knepper and M.G. Vijver, Impact of imidacloprid on daphnia magna under different food quality regimes, Environment. Toxicol. Chem., 33 (2014), 621–631. |
| [12] | R. Karimi, C. Chen, P. Pickhardt, N. Fisher and C. Folt, Stoichiometric controls of mercury dilution by growth, Proceed. Nati. Acad. Sci., 104 (2014), 7477–7482. |
| [13] | C.R. Lessard, P.C. Frost, Phosphorus nutrition alters herbicide toxicity on Daphnia magna, Sci. Total Environ., 421 (2012), 124–128. |
| [14] | I. Loladze, Y. Kuang and J.J. Elser, Stoichiometry in producer-grazer systems: Linking energy flow with element cycling, Bull Math. Bio., 62L (2000), 1137–1162. |
| [15] | D. Mergler, H.A. Anderson, L.H.M. Chan, K.R. Mahaffey, M. Murray, M. Sakamoto and A.H. Stern, Methylmercury exposure and health effects in humans: a worldwide concern, AMBIO J. Human Environ., 36 (2007),3–11. |
| [16] | F.J. Miller, P.M. Schlosser and D.B. Janszen, Habers rule: a special case in a family of curves relating concentration and duration of exposure to a fixed level of response for a given endpoint, Toxicology, 149 (2000), 21–34. |
| [17] | R.L. Morehouse, A.R. Dzialowski and P.D. Jeyasingh, Impacts of excessive dietary phosphorus on zebra mussels, Hydrobiologia, 707 (2013), 73–80. |
| [18] | A. Peace, Y. Zhao, I. Loladze, J.J. Elser and Y. Kuang, A stoichiometric producer-grazer model incorporating the effects of excess food-nutrient content on consumer dynamics, Math. Biosci., 244 (2013), 107–115. |
| [19] | A. Peace, H. Wang and Y. Kuang, Dynamics of a producer–grazer model incorporating the effects of excess food nutrient content on grazers growth, Bull. Math. Biol., 76 (2013), 2175–2197. |
| [20] | A. Peace, M. Poteat and H Wang, Somatic growth dilution of a toxicant in a predator–prey model under stoichiometric constraints, J. Theo. Biol., 407 (2013), 198–211. |
| [21] | K. Plath, M. Boersma, Mineral limitation of zooplankton: stoichiometric constraints and optimal foraging, Ecology, 82 (2013), 1260–1269. |
| [22] | R.W. Sterner and J.J. Elser, Ecological stoichiometry: the biology of elements from molecules to the biosphere, Princeton University Press. |
| [23] | M.T. Tsui, W.X. Wang, Uptake and elimination routes of inorganic mercury and methylmercury in daphnia magna, Environ. Sci. Technol., 38 (2004), 808–816. |
| [24] | R.W. Vocke, Growth responses of selected freshwater algae to trace elements and scrubber ash slurry generated by coal-fired power plants, 1978. |
| [25] | C. Walker, R. Sibly, S. Hopkin and D. Peakall, Fates of organic pollutants in individuals and organisms, Principl. Fcotoxicol., 2012 (2012), 63–93. |
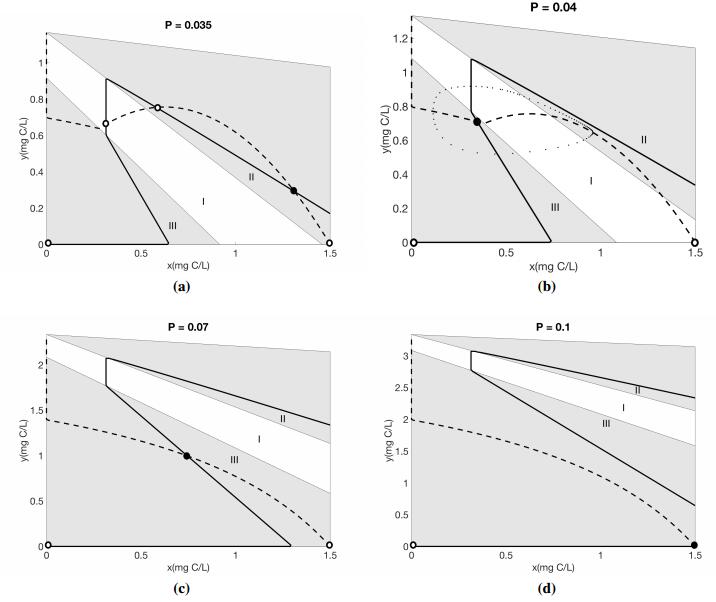
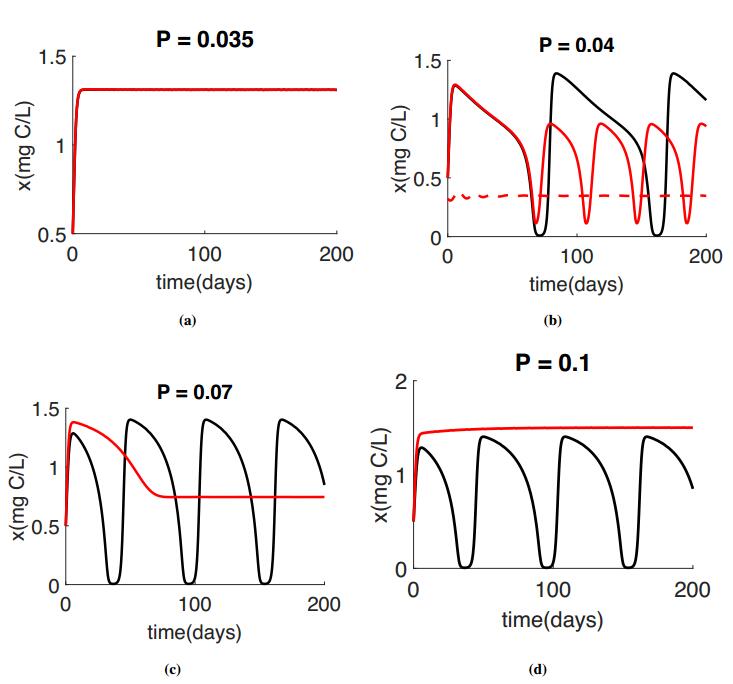
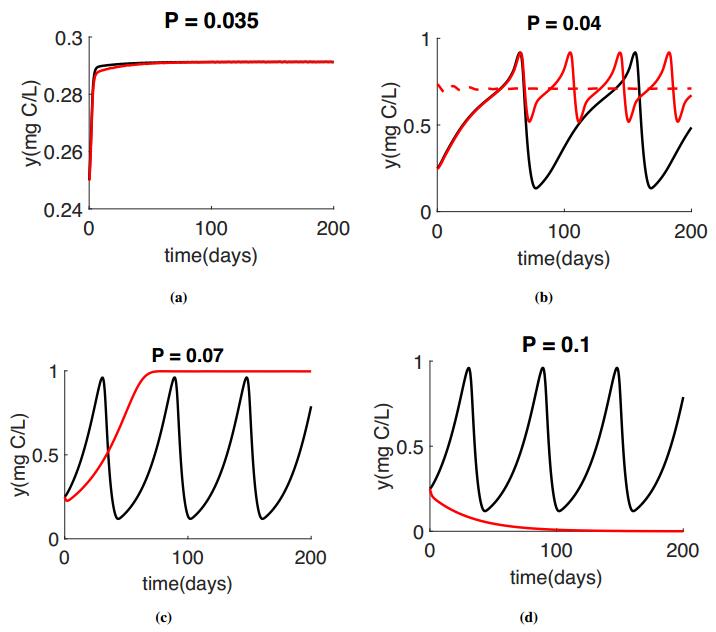
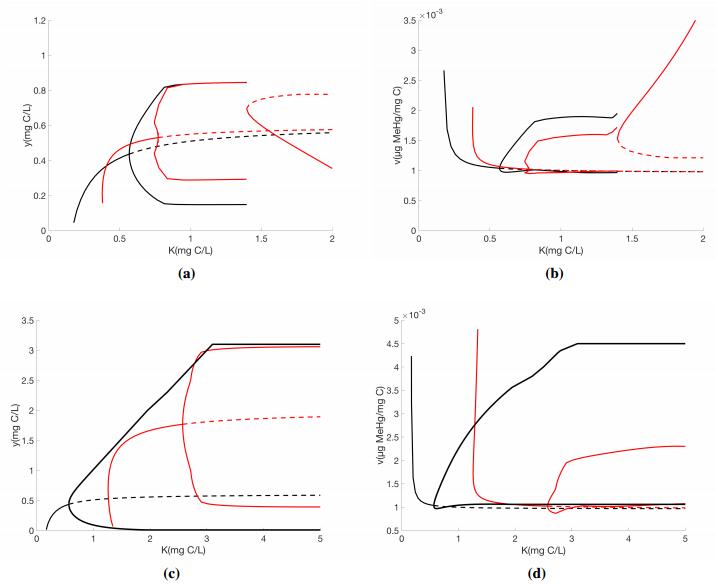
Figures(4) / Tables(1)

Md Nazmul Hassan, Kelsey Thompson, Gregory Mayer, Angela Peace. Effect of Excess Food Nutrient on Producer-Grazer Model under Stoichiometric and Toxicological Constraints[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 150-167. doi: 10.3934/mbe.2019008







 DownLoad:
DownLoad: