Citation: Patrick De Leenheer, Martin Schuster, Hal Smith. Strong cooperation or tragedy of the commons in the chemostat[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 139-149. doi: 10.3934/mbe.2019007

| [1] | K.L. Asfahl, J. Walsh, K. Gilbert and M. Schuster, Non-social adaptation defers a tragedy of the commons in Pseudomonas aeruginosa quorum sensing, ISME J., 9 (2015), 1734–1746. |
| [2] | J. S. Chuang, O. Rivoire and S. Leibler, Simpson's Paradox in a synthetic microbial system, Science, 323 (2009), 272–275. |
| [3] | J. Cremer, A. Melbinger and E. Frey, Growth dynamics and the evolution of cooperation in microbial populations, Sci Rep, 2 (2012), 281. |
| [4] | A.A. Dandekar, S. Chugani and E.P. Greenberg, Bacterial quorum sensing and metabolic incentives to cooperate, Science, 338 (2012), 264–266. |
| [5] | F. Fiegna and G.J. Velicer, Competitive fates of bacterial social parasites: persistence and selfinduced extinction of Myxococcus xanthus cheaters, Proc. Biol. Sci., 270 (2003), 1527–1534. |
| [6] | G.R. Hardin, The tragedy of the commons, Science, 162 (1968), 1243–1248. |
| [7] | R. Kümmerli, A.S. Griffin, S. A.West, A. Buckling and F. Harrison, Viscous medium promotes cooperation in the pathogenic bacterium Pseudomonas aeruginosa, Proceedings of the Royal Society B, 276 (2009), 3531–3538. |
| [8] | W.F. Lloyd, Two lectures on the checks to population (Oxford Univ. Press, Oxford, England, 1833), reprinted (in part, in: Population, Evolution, and Birth Control, G. Hardin, Ed. (Freeman, San Francisco, 1964), 37. |
| [9] | T. Pfeiffer, S. Schuster and S. Bonhoeffer, Cooperation and competition in the evolution of ATPproducing pathways, Science, 292 (2001), 504–507. |
| [10] | P.B. Rainey and K. Rainey, Evolution of cooperation and conflict in experimental bacterial populations, Nature, 425 (2003), 72–74. |
| [11] | M. Schuster, E. Foxall, D. Finch, H. Smith and P. De Leenheer, Tragedy of the commons in the chemostat, PLOS ONE, Dec 2017. Available from: https://doi.org/10.1371/journal.pone.0186119. |
| [12] | Introduction to Evolutionary Game Theory, Proceedings of Symposia in Applied Mathematics, 69 (2011), 1–25. |
| [13] | H.L. Smith, Monotone Dynamical Systems, American Mathematical Society, 1995. |
| [14] | H.L. Smith and P. Waltman, The theory of the chemostat, Cambridge University Press, 1994. |
| [15] | A.J. Waite and W. Shou, Adaptation to a new environment allows cooperators to purge cheaters stochastically, PNAS, 109 (2012), 19079–19086. |
| [16] | S.A. West, A.S. Griffin, A. Gardner and S.P. Diggle SP, Social evolution theory for microorganisms, Nat. Rev. Microbiol., 4 (2006), 597–607. |
| [17] | S.A. West, A.S. Griffin and A. Gardner, Evolutionary explanations for cooperation, Curr. Biol. 17 (2007), R661–72. |
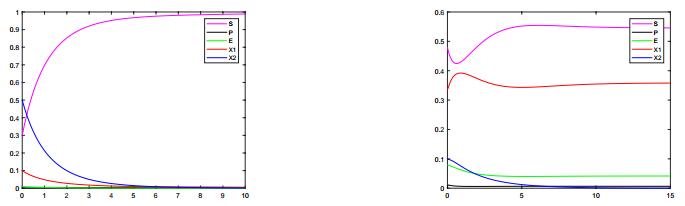
Figures(1)

Patrick De Leenheer, Martin Schuster, Hal Smith. Strong cooperation or tragedy of the commons in the chemostat[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 139-149. doi: 10.3934/mbe.2019007







 DownLoad:
DownLoad: