Citation: Ting-Hao Hsu, Tyler Meadows, LinWang, Gail S. K. Wolkowicz. Growth on two limiting essential resources in a self-cycling fermentor[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 78-100. doi: 10.3934/mbe.2019004

| [1] | D. D. Bainov and P. S. Simeonov, Systems with Impulse Effect: Stability, Theory and Applications, Ellis Horwood Series: Mathematics and its Applications, Ellis Horwood Ltd., Chichester; Halsted Press [John Wiley & Sons, Inc.], New York, 1989. |
| [2] | D. D. Bainov and P. S. Simeonov, Impulsive Differential Equations: Periodic Solutions and Applications, vol. 66 of Pitman Monographs and Surveys in Pure and Applied Mathematics, Longman Scientific & Technical, Harlow; copublished in the United States with John Wiley & Sons, Inc., New York, 1993. |
| [3] | D. D. Bainov and P. S. Simeonov, Impulsive Differential Equations: Asymptotic Properties of the Solutions, vol. 28 of Series on Advances in Mathematics for Applied Sciences, World Scientific Publishing Co., Inc., River Edge, NJ, 1995, Translated from the Bulgarian manuscript by V. Covachev [V. Khr. Kovachev]. |
| [4] | G. Butler and G. S. K. Wolkowicz, Exploitative competition in a chemostat for two complementary, and possibly inhibitory, resources, Math. Biosci., 83 (1987), 1–48. |
| [5] | A. Cinar, S. J. Parulekar, C. Ündey and G. Birol, Batch Fermentors: Modeling, Monitoring, and Control, Marcel Dekker, New York, 2003. |
| [6] | F. Córdova-Lepe, R. D. Valle and G. Robledo, Stability analysis of a self-cycling fermentation model with state-dependent impulses, Math. Methods Appl. Sci., 37 (2014), 1460–1475. |
| [7] | G. Fan and G. S. K. Wolkowicz, Analysis of a model of nutrient driven self-cycling fermentation allowing unimodal response functions, Discrete Contin. Dyn. Syst., 8 (2007), 801–831. |
| [8] | J. P. Grover, Resource Competition, Population and community biology series 19, Chapman and Hall, New York, 1997. |
| [9] | A. Halanay and D. Wexler, Teoria Calitativa a Sistemelor du Impulsuri, Ed. Acad. RSR, Bucharedt, 1968. |
| [10] | S. Hughes and D. Cooper, Biodegradation of phenol using the self-cycling fermentation process, Biotechnol. Bioeng., 51 (1996), 112–119. |
| [11] | J. A. León and D. B. Tumpson, Competition between two species for two complementary or substitutable resources, J. Theor. Biol., 50 (1975), 185–2012. |
| [12] | B. Li, G. S. K. Wolkowicz and Y. Kuang, Global asymptotic behavior of a chemostat model with two perfectly complementary resources with distributed delay, SIAM J. Appl. Math., 60 (2000), 2058–2086. |
| [13] | MATLAB, version 9.3.0 (R2017b), The MathWorks Inc., Natick, Massachusetts, 2017. |
| [14] | L. Perko, Differential Equations and Dynamical Systems, Springer-Verlag, 1996. |
| [15] | A. Samoilenko and N. Perestyuk, Impulsive Differential Equations, World Scientific, Singapore, 1995. |
| [16] | B. E. Sarkas and D. G. Cooper, Biodegradation of aromatic compounds in a self-cycling fermenter, Can. J. Chem. Eng., 72 (1994), 874–880. |
| [17] | D. Sauvageau, Z. Storms and D. G. Cooper, Sychronized populations of Escherichia coli using simplified self-cycling fermentation, J. Biotechnol., 149 (2010), 67–73. |
| [18] | H. L. Smith and P. Waltman, The Theory of the Chemostat: Dynamics of Microbial Competition., vol. 13 of Cambridge Studies in Mathematical Biology, Cambridge University Press, Cambridge, 1995. |
| [19] | R. J. Smith and G. S. K. Wolkowicz, Analysis of a model of the nutrient driven self-cycling fermentation process, Dyn. Contin. Discrete Impuls. Syst. Ser. B Appl. Algorithms, 11 (2004), 239–265. |
| [20] | Z. J. Storms, T. Brown, D. Sauvageau and D. G. Cooper, Self-cycling operation increases productivity of recombinent protein in Escherichia Coli, Biotechnol. Bioeng., 109 (2012), 2262–2270. |
| [21] | D. Tilman, Resource Competition and Community Structure, Princeton University Press, New Jersey, 1982. |
| [22] | J. Von Liebig, Die Organische Chemie in Ihrer Anwendung auf Agrikultur und Physiologie, Friedrich Vieweg, Braunschweig, 1840. |
| [23] | J. Wang, M. Chae, D. Sauvageau and D. C. Bressler, Improving ethanol productivity through selfcycling fermentation of yeast: a proof of concept, Biotechnol. Biofuels, 10 (2017), 193. |
| [24] | J. Wu, H. Nie and G. S. K. Wolkowicz, A mathematical model of competition for two essential resources in the unstirred chemostat, SIAM J. Appl. Math., 65 (2004), 209–229. |
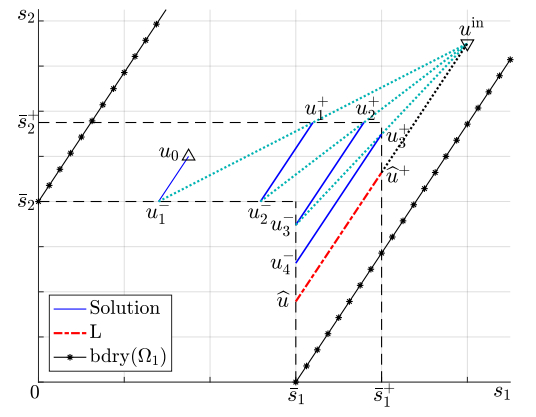
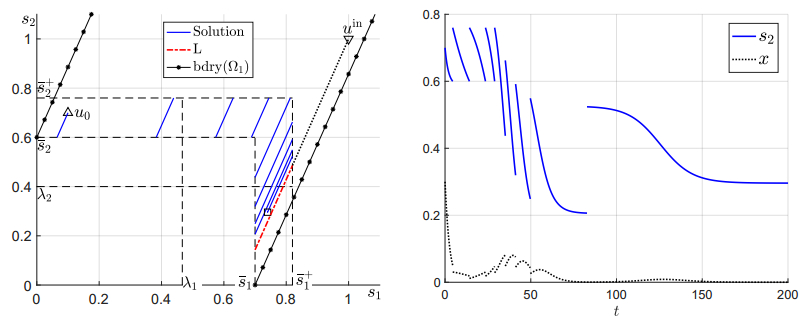
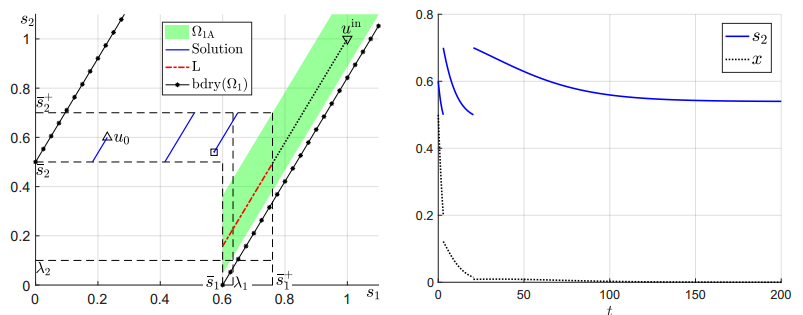
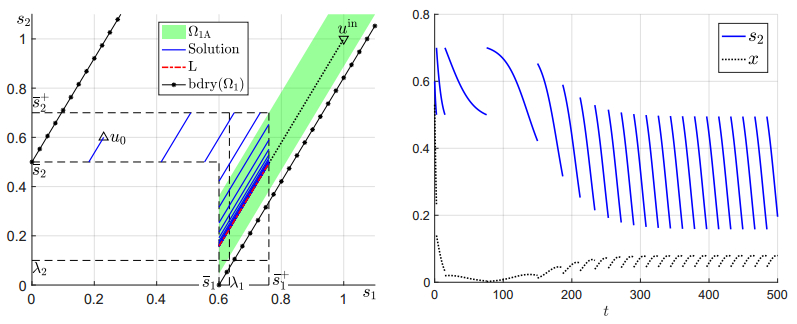
Figures(6)

Ting-Hao Hsu, Tyler Meadows, LinWang, Gail S. K. Wolkowicz. Growth on two limiting essential resources in a self-cycling fermentor[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 78-100. doi: 10.3934/mbe.2019004







 DownLoad:
DownLoad: