Citation: Xinru Zhou, Xinmiao Rong, Meng Fan, Josué-Antonio Nescolarde-Selvaa. Stoichiometric modeling of aboveground-belowground interaction of herbaceous plant[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 25-55. doi: 10.3934/mbe.2019002

| [1] | X. Y. Liu and Y. T. Mou, Research progress in the ecosystem services function and value of grassland, Acta Pratacul. Sin., 21 (2012), 286–295. |
| [2] | K. Mokany, R. Raison and A. S. Prokushkin, Critical analysis of root: shoot ratio in terrestrial biomes, Global Change Biol., 12 (2006), 84–96. |
| [3] | R. A. Gill, R. H. Kelly and W. J. Parton, Using simple environmental variables to estimate belowground productivity in grasslands, Global Biogeography, 11 (2002), 79–86. |
| [4] | J. Ares and J. S. Singh, A model of the root biomass dynamics of a shortgrass prairie dominated by blue grama. J. Appl. Ecol., 11 (1974), 727–728. |
| [5] | B. J. Enquist and K. J. Niklas, Global allocation rules for patterns of biomass partitioning in seed plants, Science, 295 (2002), 1517–1520. |
| [6] | M. Kang, C. Dai,W. Ji, Y. Jiang, Z. Yuan and H. Y. H. Chen, Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia grasslands, China, PloS One, 8 (2013), 1–7. |
| [7] | K. J. Niklas, Modelling below-and above-ground biomass for non-woody and woody plants, Ann. Bot. London, 95 (2005), 315–321. |
| [8] | W. J. Parton and D. C. Coleman, A Model of production and turnover of roots in shortgrass prairie, J. Appl. Ecol., 15 (1978), 515–541. |
| [9] | Y. Yang, J. Fang, C. Ji and W. Han, Above-and belowground biomass allocation in Tibetan grasslands, J. Veg. Sci., 20 (2009), 177–184. |
| [10] | S. Balachandra, R. J. Hull and R. A. Martins, Influence of environmental stress on biomass partitioning in transgenic tobacco plants expressing the movement protein of tobacco mosaic virus, Plant Physiol., 14 (1997), 457–481. |
| [11] | W. Fan, R. Meng and Q. S. Chen, Effects of nitrogen addition on ground/underground biomass allocation of stipa krylovii community, Anim. Husbandry Feed Sci., 31 (2010), 74–75. |
| [12] | J. H. Fank, The primary productivity of lawns in a temperate environment, J. Appl. Ecol., 17 (1980), 689–695. |
| [13] | D. E. Molyneux and W. J. Davies, Rooting pattern and water relations of three pasture grasses growing in drying soil, Oecologia, 58 (1983), 220–224. |
| [14] | X. N. Wang, The growing status and biomass partition of oriental lilies under different light intensity, Southwest China J. Agr. Sci., 20 (2007), 1091–1096. |
| [15] | W. H. Van der Putten, R. D. Bardgett, P. C. De Ruiter,W. H. G. Hol, K. M. Meyer, T. M. Bezemer, M. A. Bradford, S. Christensen, M. B. Eppinga, T. Fukami, L. Hemerik, J. Molofsky, M. Schӑdler, C. Scherber, S. Y. Strauss, M. Vos and D. A. Wardle, Empirical and theoretical challenges in aboveground-belowground ecology, Cecologia, 161 (2009), 1–14. |
| [16] | R. D. Bardgett and D. A. Wardle, Aboverground-belowground linkages: biotic interactions, ecosystem processes, and global change, Oxford University Press, 2010. |
| [17] | Z. A. Zhang and Z. Y. Chen, Plant physiology, Changchun: Jilin University Press, 2009. |
| [18] | P. Turchin, Population dynamics: a theoretical/empirical synthesis, Princeton and Oxford: Princeton university press, 2003. |
| [19] | E. P. Odem, Energy flow in ecosystems: a historical view, Amer. Zool, 8 (1968), 11–18. |
| [20] | W. A. Reiners, Complementary models for ecosystems, Am. Nat., 127 (1986), 59–73. |
| [21] | N. Loeuille, M. Loreau and R. Ferrière, Consequences of plant-herbivore coevolution on the dynamics and function of ecosystems, J. Theor. Biol., 217 (2002), 369–381. |
| [22] | J. J. Elser, R. W. Sterner and E. Gorokhova, Biological stoichiometry from genes to ecosystems, Ecol. Lett., 3 (2000), 540–550. |
| [23] | M. Fan, I. Loladze and Y. Kuang, Dynamics of a stoichiometric discrete producer-grazer model, J. Differ. Equ. Appl., 11 (2005), 347–364. |
| [24] | X. Li, H. Wang and Y. Kuang, Global analysis of a stoichiometry producer-grazer model with holling type functional responses, J. Math. Biol., 63 (2011), 901–932. |
| [25] | I. Loladez and Y. Kuang, Stoichiometry in producer-grazer system: linking energy flow with element cycling, B. Math. Biol., 62 (2000), 1137–1162. |
| [26] | I. Loladze, Y. Kuang and J. Elser, Competition and stoichiometry: coexistence of two predators on one prey, Theor. Popul. Biol., 65 (2004), 1–15. |
| [27] | H. Wang, Y. Kuang and I. Loladze, Dynamics of a mechanistically derived stoichiometric producer-grazer model, J. Biol. Dynam., 2 (2008), 286–296. |
| [28] | W. T. Liu, Z. J. Wei, S. J. Lu and S. X. Sun, Research advances in stoichiometry of grassland in China, Acta Agrestia Sinica, 5 (2015), 916–926. |
| [29] | J. Penuelas and I. Filella, Herbaria century record of increasing eutrophication in Spanish terrestrial ecosystem, Global Change Biol., 7 (2011), 427–433. |
| [30] | S. A. L. M. Kooijman, The stoichiometry of animal energetics, J. Thero. Biol., 177 (1995), 139–149. |
| [31] | D. R. Bouldin, Mathematical description of diffusion process in the soil-plant system, Soil Sci. Soc. Am., 25 (1961), 476–480. |
| [32] | X. Z. Lv, Z. Yu and G. Y. Wang, Element biology, Hefei: University of Science and Technology China press, (2011), 199–201. |
| [33] | E. B. Muller, R. M. Nisbet, S. A. Kooijman, J. J. Elser and E. McCauley, Stoichiometric food quality and herbivore dynamics, Ecol. Lett., 4 (2001), 519–529. |
| [34] | X. Y. Ping, B. R. Jia, W. P. Yuan, F. Y. Wang, Y. H. Wang and L. Zhou, Biomass allocation of leymus chinensis population: a dynamic simulation study, Chinese J. Appl. Ecol., 18 (2007), 2699–2704. |
| [35] | C. J. Li, Advanced plant nutrition, Beijing: China Agricultural University press, 2008. |
| [36] | C. Ma, The influence on different nitrogen level to the nitrogen absorption rate in soybean, Soybean Science, 24 (2005), 52–63. |
| [37] | S. E. Jørgensen and G. Bendoricchio, Fundamentals of ecological modelling, Elsevier, 2001. |
| [38] | Z. J. Bin, J. J.Wang andW. P. Zhang, Effects of n addition on ecological stoichiometric characteristics in six dominant species of alpine meadow on the Qinghai-Tibetan Plateau, China, J. Plant Ecol., 38 (2014), 231–237. |
| [39] | X. J. Xin, Effects of N,P addition on above/below-ground biomass allocation and plant functional type's composition in a sub-alpine meadow, Ecological Science, 33 (2014), 452–458. |
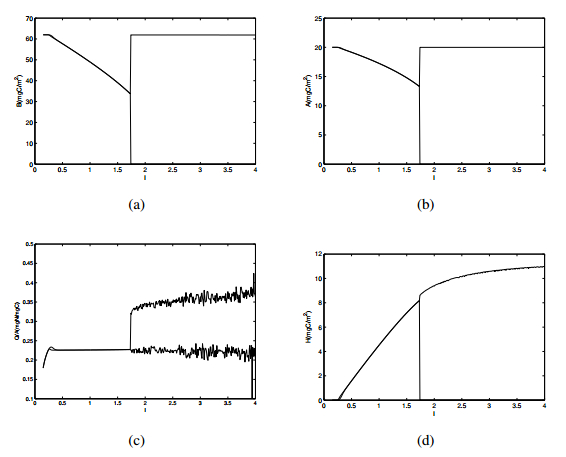
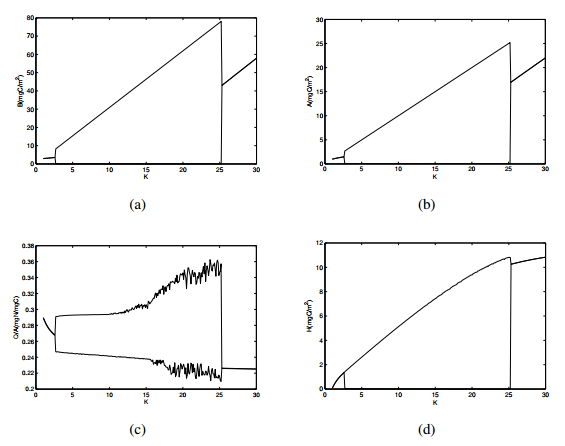
Figures(10) / Tables(1)

Xinru Zhou, Xinmiao Rong, Meng Fan, Josué-Antonio Nescolarde-Selvaa. Stoichiometric modeling of aboveground-belowground interaction of herbaceous plant[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 25-55. doi: 10.3934/mbe.2019002







 DownLoad:
DownLoad: