Citation: Andrea Pugliese, Abba B. Gumel, Fabio A. Milner, Jorge X. Velasco-Hernandez. Sex-biased prevalence in infections with heterosexual, direct, and vector-mediated transmission: a theoretical analysis[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 125-140. doi: 10.3934/mbe.2018005

| [1] | [ C. L. Althaus,N. Low, How Relevant Is Sexual Transmission of Zika Virus?, PLOS Med., 13 (2016): 1-3. |
| [2] | [ M.-C. Boily,R. F. Baggaley,L. Wang,B. Masse,R. G. White,R. J. Hayes,M. Alary, Heterosexual risk of HIV-1 infection per sexual act: Systematic review and meta-analysis of observational studies, Lancet Infect. Dis., 9 (2009): 118-129. |
| [3] | [ R. B. Brooks,M. P. Carlos,R. A. Myers,M. Grace White,T. Bobo-Lenoci,D. Aplan,D. Blythe,K. A. Feldman, Likely Sexual Transmission of Zika Virus from a Man with No Symptoms of Infection --Maryland, 2016, MMWR Morb Mortal Wkly Rep, 65 (2016): 915-916. |
| [4] | [ CDC, Hepatitis A Questions and Answers for the Public, URL https://www.cdc.gov/hepatitis/hav/afaq.htm. |
| [5] | [ F. C. Coelho,B. Durovni,V. Saraceni,C. Lemos,C. T. Codeco,S. Camargo,L. M. de Carvalho,L. Bastos,D. Arduini,D. A. M. Villela,M. Armstrong, Higher incidence of Zika in adult women than adult men in Rio de Janeiro suggests a significant contribution of sexual transmission from men to women, Int. J. Infect. Dis., 51 (2016): 128-132. |
| [6] | [ W. D. Davidson,S Slavinski,K. Komoto,J. Rakeman, Suspected Female-to-Male Sexual Transmission of Zika Virus-New York City, 2016, MMWR Morb Mortal Wkly Rep, 65 (2016): 716-717. |
| [7] | [ O. Diekmann and J. A. P. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, New York: John Wiley and Sons, 2000. |
| [8] | [ M. R. Duffy,T.-H. Chen,W. T. Hancock,A. M. Powers,J. L. Kool,R. S. Lanciotti,M. Pretrick,M. Marfel,S. Holzbauer,C. Dubray,L. Guillaumot,A. Griggs,M. Bel,A. J. Lambert,J. Laven,O. Kosoy,A. Panella,B. J. Biggerstaff,M. Fischer,E. B. Hayes, Zika virus outbreak on Yap Island, Federated States of Micronesia., N. Engl. J. Med., 360 (2009): 2536-2543. |
| [9] | [ N. M. Ferguson, Z. M. Cucunubá, I. Dorigatti, G. L. Nedjati-Gilani, C. A. Donnelly, M.- G. Basáñez, P. Nouvellet and J. Lessler, Countering the Zika epidemic in Latin America, Science (80-. )., 353 (2016), 353-354, URL http://www.sciencemag.org/cgi/content/full/science.aag0219/DC1. |
| [10] | [ K. Fonseca, B. Meatherall, D. Zarra, M. Drebot, J. MacDonald, K. Pabbaraju, S. Wong, P.Webster, R. Lindsay and R. Tellier, First case of zika virus infection in a returning canadian traveler, Am. J. Trop. Med. Hyg., 91 (2014), 1035-1038, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4228871/. |
| [11] | [ P. J. Fos, null, Epidemiology Foundations: The Science of Public Health, Jossey-Bass, San Francisco, 2011. |
| [12] | [ E. B. Hayes, Zika Virus Outside Africa, Emerg. Infect. Dis., 15 (2009), 1347-1350, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2819875/http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2819875/. |
| [13] | [ W. O. Kermack,A. G. McKendrick, A contributions to the mathematical theory of epidemics, Proc. R. Soc. London A, 115 (1927): 700-721. |
| [14] | [ A. J. Kucharski,S. Funk,R. M. Eggo,H.-P. Mallet,W. J. Edmunds,E. J. Nilles, Transmission dynamics of zika virus in island populations: A modelling analysis of the 2013-14 french polynesia outbreak, PLoS Negl. Trop. Dis, 10 (2016): e0004726. |
| [15] | [ W. D. Z. Lopes, J. D. Rodriguez, F. A. Souza, T. R. dos Santos, R. S. dos Santos, W. M. Rosanese, W. R. Z. Lopes, C. A. Sakamoto and A. J. da Costa, Sexual transmission of Toxoplasma gondii in sheep, Vet. Parasitol., 195 (2013), 47-56, URL http://linkinghub.elsevier.com/retrieve/pii/S0304401713000083. |
| [16] | [ G. MacDonald, The analysis of equilibrium in malaria, Trop Dis Bull, 49 (1952): 813-829. |
| [17] | [ J. M. Mansuy,C. Pasquier,M. Daudin,S. Chapuy-Regaud,N. Moinard,C. Chevreau,J. Izopet,C. Mengelle,L. Bujan, Zika virus in semen of a patient returning from a non-epidemic area, Lancet Infect. Dis., 16 (2016): 894-895. |
| [18] | [ J. C. Miller, Mathematical models of SIR disease spread with combined non-sexual and sexual transmission routes, Infect. Dis. Model., 2 (2017), 35-55, URL http://linkinghub.elsevier.com/retrieve/pii/S2468042716300203. |
| [19] | [ R. Pellissier,A. Rousselot, Enquete serologique sur l'incidence des virus neurol ropes chez quelques singes de l'Afrique Equatoriale Francaise (French) [A Serological Investigation of the Incidence of Neurotropic Viruses in Certain Monkeys of French Equatorial Africa], Bull. Société Pathol. Exot., 47 (1954): 228-231. |
| [20] | [ A. J. Rodriguez-Morales, A. C. Bandeira and C. Franco-Paredes, The expanding spectrum of modes of transmission of Zika virus: A global concern, Ann. Clin. Microbiol. Antimicrob. , 15 (2016), 13, URL http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4776405/. |
| [21] | [ R. Ross, The Prevention of Malaria, Churchill, London, 1911. |
| [22] | [ Q. Zhang, K. Sun, M. Chinazzi, A. Pastore-Piontti, N. E. Dean, D. P. Rojas, S. Merler, D. Mistry, P. Poletti, L. Rossi, M. Bray, M. E. Halloran, I. M. Longini and A. Vespignani, Spread of Zika virus in the Americas, Proc. Natl. Acad. Sci. (2017). |
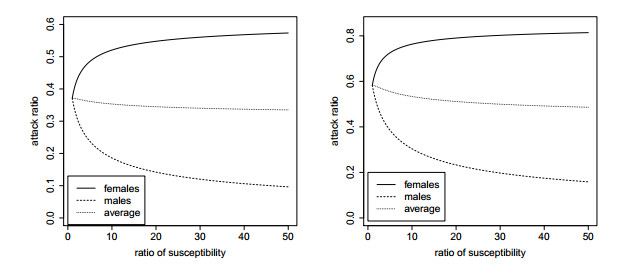
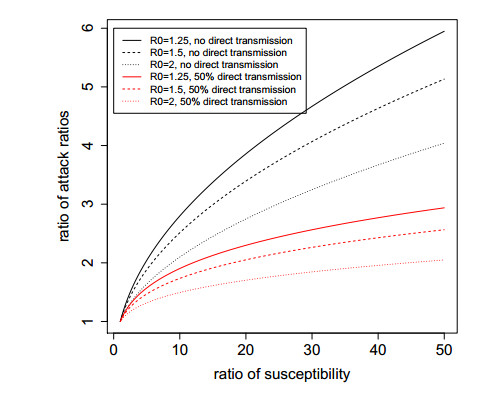
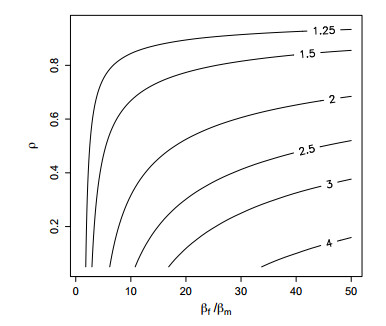
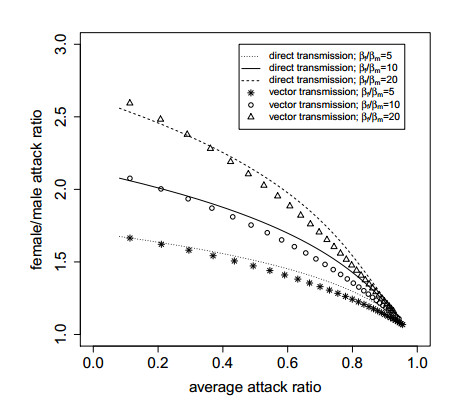
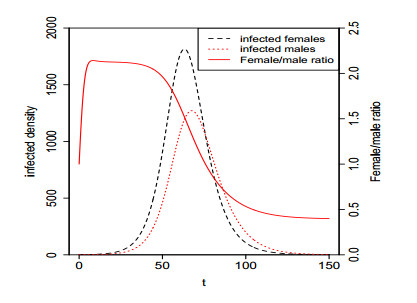
Figures(5)

Andrea Pugliese, Abba B. Gumel, Fabio A. Milner, Jorge X. Velasco-Hernandez. Sex-biased prevalence in infections with heterosexual, direct, and vector-mediated transmission: a theoretical analysis[J]. Mathematical Biosciences and Engineering, 2018, 15(1): 125-140. doi: 10.3934/mbe.2018005







 DownLoad:
DownLoad: