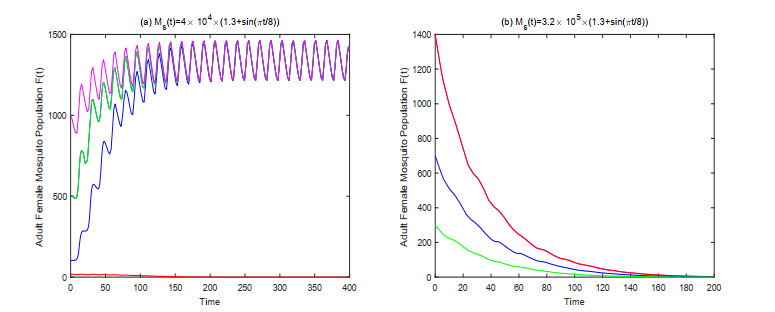
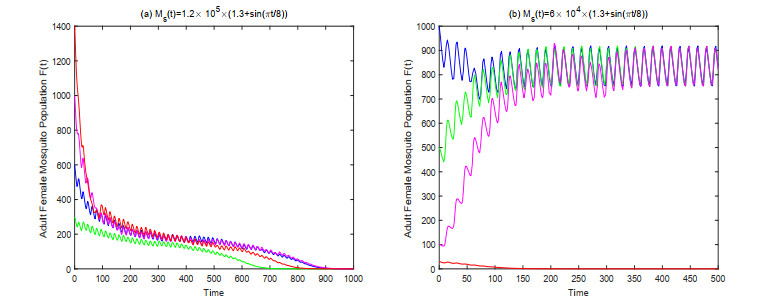
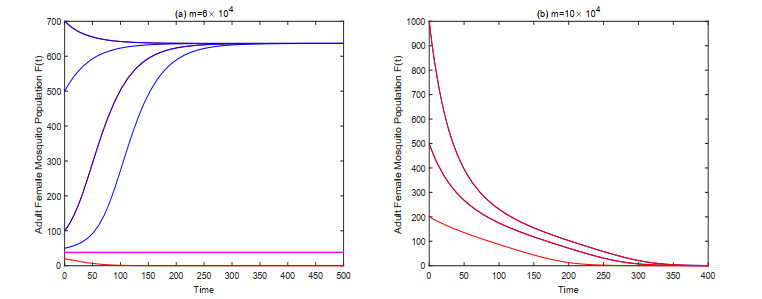
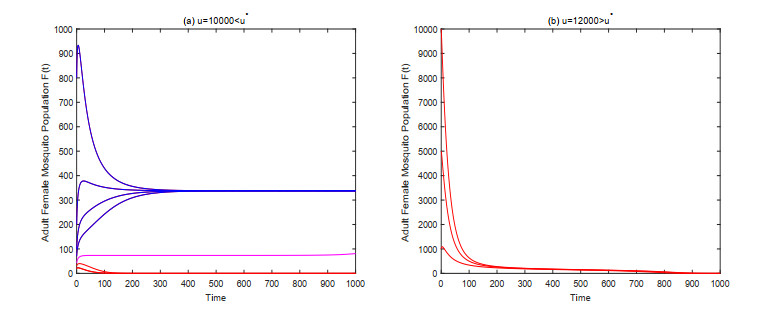
In this paper, we study a kind of mosquito population suppression model incorporating the growth stage as well as the sex structure of mosquitoes. For the general non-autonomous case, a threshold $ m^* $ for the number of sexually active sterile mosquitoes in the field is defined, and sufficient conditions for successful suppression and partial suppression of wild mosquito population are obtained. For the first special case when the release period of the sterile mosquito is equal to its sexual lifespan, we determined the initial population size under which the mosquito population can be suppressed finally for different release intensities. For the latter special case when sterile mosquitoes are released with a constant rate, we give a threshold $ u^* $ for the release rate, and investigate the dynamic behavior of the system based on this threshold. Finally, some numerical examples are presented to confirm the theoretical results.
Citation: Junjie He, Di Li, Shouzong Liu. Global dynamics of a mosquito population suppression model with stage and sex structure[J]. AIMS Mathematics, 2023, 8(6): 14027-14046. doi: 10.3934/math.2023717
In this paper, we study a kind of mosquito population suppression model incorporating the growth stage as well as the sex structure of mosquitoes. For the general non-autonomous case, a threshold $ m^* $ for the number of sexually active sterile mosquitoes in the field is defined, and sufficient conditions for successful suppression and partial suppression of wild mosquito population are obtained. For the first special case when the release period of the sterile mosquito is equal to its sexual lifespan, we determined the initial population size under which the mosquito population can be suppressed finally for different release intensities. For the latter special case when sterile mosquitoes are released with a constant rate, we give a threshold $ u^* $ for the release rate, and investigate the dynamic behavior of the system based on this threshold. Finally, some numerical examples are presented to confirm the theoretical results.

| [1] |
S. Ai, J. Li, J. Yu, B. Zheng, Stage-structured models for interactive wild and periodically and impulsively released sterile mosquitoes, Discrete Cont. Dyn-B, 27 (2022), 3039–3052. https://doi.org/10.3934/dcdsb.2021172 doi: 10.3934/dcdsb.2021172

|
| [2] |
L. Almeida, M. Duprez, Y. Privat, N. Vauchelet, Optimal control strategies for the sterile mosquitoes technique, J. Differ. Equations, 311 (2022) 229–266. https://doi.org/10.1016/j.jde.2021.12.002 doi: 10.1016/j.jde.2021.12.002
|
| [3] |
L. Almeida, M. Duprez, Y. Privat, N. Vauchelet, Mosquito population control strategies for fighting against arboviruses, Math. Biosci. Eng., 16 (2019) 6274–6297. https://doi.org/10.3934/mbe.2019313 doi: 10.3934/mbe.2019313
|
| [4] |
R. Anguelov, Y. Dumont, I. Djeumen, Sustainable vector/pest control using the permanent Sterile Insect Technique, Math. Method. Appl. Sci., 43 (2020), 10391–10412. https://doi.org/10.1002/mma.6385 doi: 10.1002/mma.6385
|
| [5] |
H. J. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol., 24 (1982), 405–416. https://doi.org/10.1007/BF02515585 doi: 10.1007/BF02515585
|
| [6] |
H. J. Barclay, M. Mackuer, The sterile insect release method for pest control: a density dependent model, Environ. Entomol., 9 (1980), 810–817. https://doi.org/10.1093/ee/9.6.810 doi: 10.1093/ee/9.6.810
|
| [7] |
P. A. Bliman, D. Cardona-Salgado, Y. Dumont, O. Vasilieva, Implementation of control strategies for sterile insect techniques, Math. Biosci., 314 (2019), 43–60. https://doi.org/10.1016/j.mbs.2019.06.002 doi: 10.1016/j.mbs.2019.06.002
|
| [8] |
G. Briggs, The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti, PLoS Pathog., 6 (2010), e1000833. https://doi.org/10.1371/journal.ppat.1000833 doi: 10.1371/journal.ppat.1000833
|
| [9] |
L. Cai, S. Ai, J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM, J. Appl. Math., 74 (2014), 1786–1809. https://doi.org/10.1137/13094102X doi: 10.1137/13094102X
|
| [10] |
F. Gazori, M. Hesaaraki, Three-dimensional spread analysis of a Dengue disease model with numerical season control, Int. J. Biomath., 14 (2021), 2150066. https://doi.org/10.1142/S1793524521500662 doi: 10.1142/S1793524521500662
|
| [11] |
M. Guo, L. Hu, L. Nie, Stochastic dynamics of the transmission of Dengue fever virus between mosquitoes and humans, Int. J. Biomath., 14 (2021), 2150062. https://doi.org/10.1142/S1793524521500625 doi: 10.1142/S1793524521500625
|
| [12] |
M. Huang, J. Luo, L. Hu, B. Zheng, J. Yu, Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theor. Biol., 440 (2018), 1–11. https://doi.org/10.1016/j.jtbi.2017.12.012 doi: 10.1016/j.jtbi.2017.12.012
|
| [13] |
M. Huang, S. Liu, X. Song, Study of the sterile insect release technique for a two-sex mosquito population model, Math. Biosci. Eng., 18 (2021), 1314–1339. https://doi.org/10.3934/mbe.2021069 doi: 10.3934/mbe.2021069
|
| [14] |
M. Huang, L. You, S. Liu, X. Song, Impulsive release strategies of sterile mosquitos for optimal control of wild population, J. Biol. Dynam., 15 (2021), 151–176. https://doi.org/10.1080/17513758.2021.1887380 doi: 10.1080/17513758.2021.1887380
|
| [15] |
M. Huang, X. Song, J. Li, Modelling and analysis of impulsive release of sterile mosquitoes, J. Biol. Dynam., 11 (2017), 147–171. https://doi.org/10.1080/17513758.2016.1254286 doi: 10.1080/17513758.2016.1254286
|
| [16] |
M. Huang, S. Liu, X. Song, Study of a delayed mosquito population suppression model with stage and sex structure, J. Appl. Math. Comput., 69 (2023), 89–111. https://doi.org/10.1007/s12190-022-01735-w doi: 10.1007/s12190-022-01735-w
|
| [17] |
Y. Hui, Z. Zhao, Q. Li, L. Pang, Asymptotic stability in a mosquito population suppression model with time delay, Int. J. Biomath., 16 (2023), 2250092. https://doi.org/10.1142/S1793524522500929 doi: 10.1142/S1793524522500929
|
| [18] |
J. Li, S. Ai, Impulsive releases of sterile mosquitoes and interactive dynamics with time delay, J. Biol. Dynam., 14 (2020), 313–331. https://doi.org/10.1080/17513758.2020.1748239 doi: 10.1080/17513758.2020.1748239
|
| [19] |
J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dynam., 11 (2017), 316–333. https://doi.org/10.1080/17513758.2016.1216613 doi: 10.1080/17513758.2016.1216613
|
| [20] |
G. Lin, Y. Hui, Stability analysis in a mosquito population suppression model, J. Biol. Dyn., 14 (2020), 578–589. https://doi.org/10.1080/17513758.2020.1792565 doi: 10.1080/17513758.2020.1792565
|
| [21] |
A. Lupica, A. Palumbo, The coexistence of fast and slow diffusion processes in the life cycle of Aedes aegypti mosquitoes, Int. J. Biomath., 14 (2021), 2050087. https://doi.org/10.1142/S1793524520500874 doi: 10.1142/S1793524520500874
|
| [22] |
Y. Pang, S. Wang, S. Liu, Dynamics analysis of stage-structured wild and sterile mosquito interaction impulsive model, J. Biol. Dyn., 16 (2022), 464–479. https://doi.org/10.1080/17513758.2022.2079739 doi: 10.1080/17513758.2022.2079739
|
| [23] |
M. Strugarek, H. Bossin, Y. Dumont, On the use of the sterile insect release technique to reduce or eliminate mosquito populations, Appl. Math. Model., 68 (2019), 443–470. https://doi.org/10.1016/j.apm.2018.11.026 doi: 10.1016/j.apm.2018.11.026
|
| [24] |
J. Yu, Modeling Mosquito Population Suppression Based on Delay Differential Equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. https://doi.org/10.1137/18M1204917 doi: 10.1137/18M1204917
|
| [25] |
J. Yu, J. Li, Global asymptotic stability in an interactive wild and sterile mosquito model, J. Differ. Equations, 269 (2020), 6193–6215. https://doi.org/10.1016/j.jde.2020.04.036 doi: 10.1016/j.jde.2020.04.036
|
| [26] |
J. Yu, Existence and stability of a unique and exact two periodic orbits for an interactive wild and sterile mosquito model, J. Differ. Equations, 269 (2020), 10395–10415. https://doi.org/10.1016/j.jde.2020.07.019 doi: 10.1016/j.jde.2020.07.019
|
| [27] |
J. Yu, J. Li, Dynamics of interactive wild and sterile mosquitoes with time delay, J. Biol. Dynam., 13 (2019), 606–620. https://doi.org/10.1080/17513758.2019.1682201 doi: 10.1080/17513758.2019.1682201
|
| [28] |
M. Zhang, Z. Lin, H. Zhu, The transmission of dengue virus with Aedes aegypti mosquito in a heterogeneous environment, Int. J. Biomath., 14 (2021), 2150026. https://doi.org/10.1142/S1793524521500261 doi: 10.1142/S1793524521500261
|
| [29] | Z. Zhao, L. Pang, X. Song, D. Wang, Q. Li, Impact of the impulsive releases and Allee effect on the dispersal behavior of the wild mosquitoes, J. Appl. Math. Comput., 68 (2022), 1527–1544. |
| [30] |
X. Zheng, D. Zhang, Y. Li, C. Yang, Y. Wu, X. Liang, et al., Incompatible and sterile insect techniques combined eliminate mosquitoes, Nature, 572 (2019), 56–61. https://doi.org/10.1038/s41586-019-1407-9 doi: 10.1038/s41586-019-1407-9
|
| [31] |
B. Zheng, J. Yu, J. Li, Existence and stability of periodic solutions in a mosquito population suppression model with time delay, J. Differ. Equations, 315 (2022), 159–178. https://doi.org/10.1016/j.jde.2022.01.036 doi: 10.1016/j.jde.2022.01.036
|
| [32] |
B. Zheng, J. Yu, J. Li, Modeling and analysis of the implementation of the Wolbachia incompatible and sterile insect technique for mosquito population suppression, SIAM J. Appl. Math., 81 (2021), 718–740. https://doi.org/10.1137/20M1368367 doi: 10.1137/20M1368367
|
| [33] |
B. Zheng, J. Yu, Existence and uniqueness of periodic orbits in a discrete model on Wolbachia infection frequency, Adv. Nonlinear Anal., 11 (2022), 212–224. https://doi.org/10.1515/anona-2020-0194 doi: 10.1515/anona-2020-0194
|
| [34] |
B. Zheng, J. Li, J. Yu, One discrete dynamical model on Wolbachia infection frequency in mosquito populations, Sci. China Math., 65 (2022), 1749–1764. https://doi.org/10.1007/s11425-021-1891-7 doi: 10.1007/s11425-021-1891-7
|
| [35] |
Z. Zhu, B. Zheng, Y. Shi, R. Yan, J. Yu, Stability and periodicity in a mosquito population suppression model composed of two sub-models, Nonlinear Dynam., 107 (2022), 1383–1395. https://doi.org/10.1007/s11071-021-07063-1 doi: 10.1007/s11071-021-07063-1
|
Figures(5) / Tables(2)

Junjie He, Di Li, Shouzong Liu. Global dynamics of a mosquito population suppression model with stage and sex structure[J]. AIMS Mathematics, 2023, 8(6): 14027-14046. doi: 10.3934/math.2023717







 DownLoad:
DownLoad: