In this article, we deal with the following fractional p-Kirchhoff type equation
{M(∫RN∫RN|u(x)−u(y)|p|x−y|N+psdxdy)(−Δ)spu=|u|p∗α−2u|x|α+λ|x|β,in Ω,u>0,in Ω,u=0,in RN∖Ω,
where Ω⊂RN is a smooth bounded domain containing 0, (−Δ)sp denotes the fractional p-Laplacian, M(t)=a+btk−1 for t≥0 and k>1, a,b>0, λ>0 is a parameter, 0<s<1, 0≤α<ps<N, N(p−2)+psp−1<β<N(p∗α−1)+αp∗α, 1<p<pk<p∗α=p(N−α)N−ps is the fractional critical Hardy-Sobolev exponent. With aid of the variational method and the concentration compactness principle, we prove the existence of two distinct positive solutions.
Citation: Zusheng Chen, Hongmin Suo, Jun Lei. Multiple solutions for a fractional p-Kirchhoff equation with critical growth and low order perturbations[J]. AIMS Mathematics, 2022, 7(7): 12897-12912. doi: 10.3934/math.2022714
| [1] | Zhaoping Qin, Patrick Robichaud, Taihao Quan . Oxidative stress and CCN1 protein in human skin connective tissue aging. AIMS Molecular Science, 2016, 3(2): 269-279. doi: 10.3934/molsci.2016.2.269 |
| [2] | Akshaj Pole, Manjari Dimri, Goberdhan P. Dimri . Oxidative stress, cellular senescence and ageing. AIMS Molecular Science, 2016, 3(3): 300-324. doi: 10.3934/molsci.2016.3.300 |
| [3] | Noor Riyadh Thiab, Nicola King, Mary McMillan, Amer Almashhadany, Graham L Jones . Age-related protein and mRNA expression of glutathione peroxidases (GPx) and Hsp-70 in different regions of rat kidney with and without stressor. AIMS Molecular Science, 2016, 3(2): 125-137. doi: 10.3934/molsci.2016.2.125 |
| [4] | Sayeeda Ahsanuddin, Minh Lam, Elma D. Baron . Skin aging and oxidative stress. AIMS Molecular Science, 2016, 3(2): 187-195. doi: 10.3934/molsci.2016.2.187 |
| [5] | Mariacristina De Luca, Kevin Pels, Susana Moleirinho, Graziella Curtale . The epigenetic landscape of innate immunity. AIMS Molecular Science, 2017, 4(1): 110-139. doi: 10.3934/molsci.2017.1.110 |
| [6] | Brandt D. Pence . Recent developments and future perspectives in aging and macrophage immunometabolism. AIMS Molecular Science, 2021, 8(3): 193-201. doi: 10.3934/molsci.2021015 |
| [7] | Giulia Ambrosi, Pamela Milani . Endoplasmic reticulum, oxidative stress and their complex crosstalk in neurodegeneration: proteostasis, signaling pathways and molecular chaperones. AIMS Molecular Science, 2017, 4(4): 424-444. doi: 10.3934/molsci.2017.4.424 |
| [8] | Dora Brites . Cell ageing: a flourishing field for neurodegenerative diseases. AIMS Molecular Science, 2015, 2(3): 225-258. doi: 10.3934/molsci.2015.3.225 |
| [9] | Michael W Patt, Lisa Conte, Mary Blaha, Balbina J Plotkin . Steroid hormones as interkingdom signaling molecules: Innate immune function and microbial colonization modulation. AIMS Molecular Science, 2018, 5(1): 117-130. doi: 10.3934/molsci.2018.1.117 |
| [10] | Hannah P. Priyanka, Rahul S. Nair, Sanjana Kumaraguru, Kirtikesav Saravanaraj, Vasantharekha Ramasamy . Insights on neuroendocrine regulation of immune mediators in female reproductive aging and cancer. AIMS Molecular Science, 2021, 8(2): 127-148. doi: 10.3934/molsci.2021010 |
In this article, we deal with the following fractional p-Kirchhoff type equation
{M(∫RN∫RN|u(x)−u(y)|p|x−y|N+psdxdy)(−Δ)spu=|u|p∗α−2u|x|α+λ|x|β,in Ω,u>0,in Ω,u=0,in RN∖Ω,
where Ω⊂RN is a smooth bounded domain containing 0, (−Δ)sp denotes the fractional p-Laplacian, M(t)=a+btk−1 for t≥0 and k>1, a,b>0, λ>0 is a parameter, 0<s<1, 0≤α<ps<N, N(p−2)+psp−1<β<N(p∗α−1)+αp∗α, 1<p<pk<p∗α=p(N−α)N−ps is the fractional critical Hardy-Sobolev exponent. With aid of the variational method and the concentration compactness principle, we prove the existence of two distinct positive solutions.
Age-related macular degeneration (AMD) is a well-established disease of aging and chronic inflammation. The hallmark of the disease condition is the presence of soft drusen, yellow deposits of lipids and proteins, primarily in the area of the central region of retina called macula (Figure 1). However, the presence of confluent, soft drusen is not sufficient for the clinical diagnosis of AMD. Rather, at this early/intermediate stage of AMD tissue loss is either absent or only in its early stages and vision is usually unaffected [1]. AMD progression and severity is directly correlated to the number and size of drusen. Advanced AMD occurs in 2 forms: (i) geographic atrophy (GA) of the retinal pigment epithelium (RPE) and overlying photoreceptors (also called advanced “dry” AMD), and (ii) choroidal neovascularization (CNV, also called “wet” AMD). Dry (GA) AMD is characterized by confluent areas of photoreceptor and RPE cell death, and is responsible for 10% of the legal blindness caused by AMD [2]. Approximately 900,000 people in the United States are currently affected by GA with more than half of the patients occurring bilaterally [3]. Wet (CNV) AMD accounts for the remaining 90% of acute blindness caused by AMD and is characterized by abnormal blood vessel growth under the macula. These new vessels are largely malformed, which leads to the improper vascular integrity causing undesirable fluid leakage within the disrupted tissue infiltrated by the unwanted vasculature (Figure 2) [4,5].
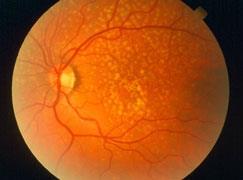 Figure 1. Drusen are yellow deposits under the retina, the light-sensitive tissue at the back of the eye. Drusen consist of lipids and fatty protein. While not all drusen cause AMD, their presence increases a person’s risk of developing AMD. (Adapted from [7]).
Figure 1. Drusen are yellow deposits under the retina, the light-sensitive tissue at the back of the eye. Drusen consist of lipids and fatty protein. While not all drusen cause AMD, their presence increases a person’s risk of developing AMD. (Adapted from [7]).
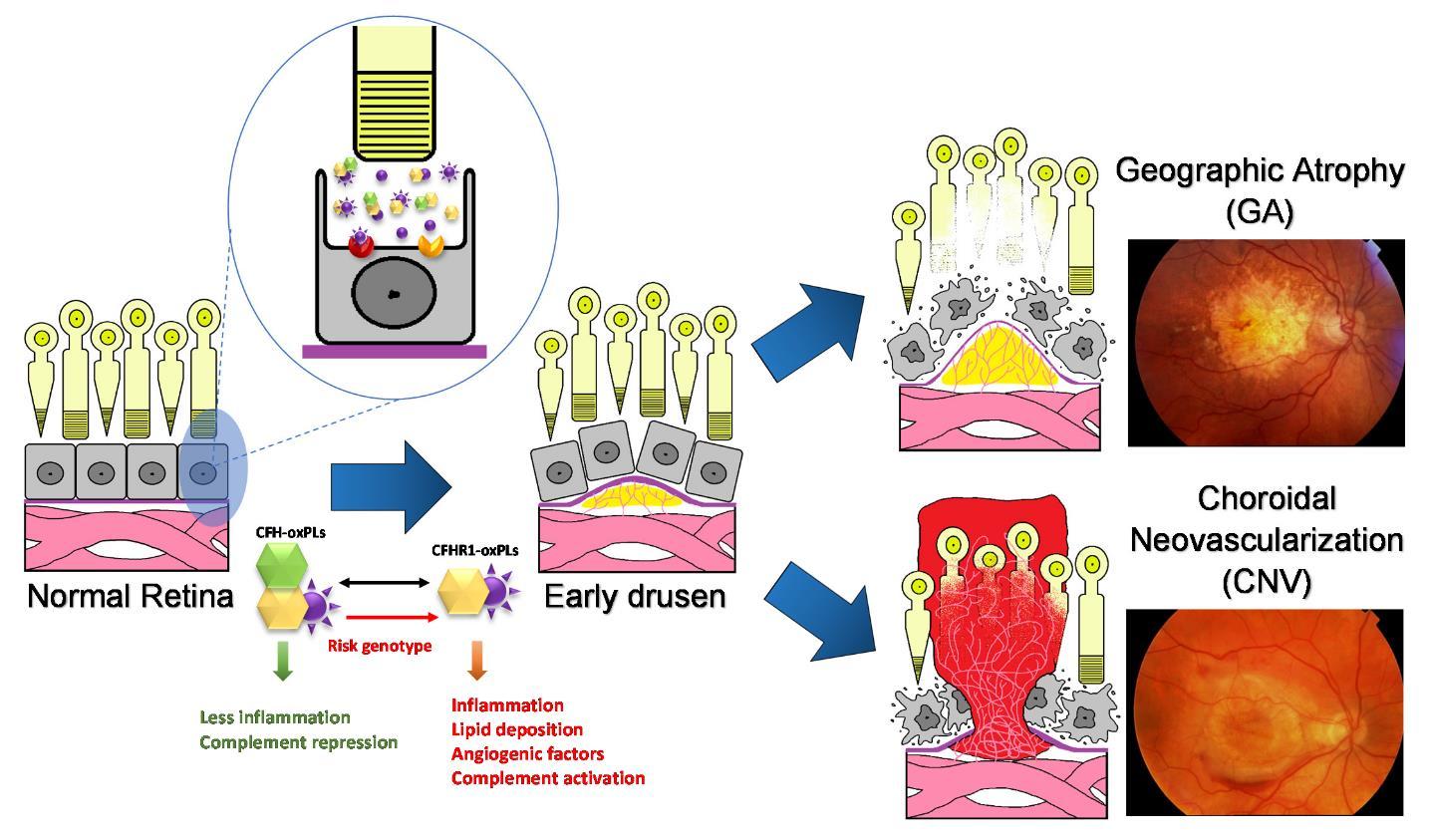 Figure 2. Age-related macular degeneration (AMD) is a disease that causes the progressive damage of the macula, the center of retina responsive for central and precise vision.Genetic risk factors response to the environment stimulants, such as oxidative stress resulting in drusen formation, inflammation and abnormal vascular growth. The unique nature of the eye leads to an abnormal burded of both degraded lipid products and oxidative stress, leading to relatively greater burden of oxidized lipid biproducts such as oxPLs. There are two forms of advanced AMD: graphic atrophy (GA) (upper right) and choroidal neovascularization (CNV) (lower right).
Figure 2. Age-related macular degeneration (AMD) is a disease that causes the progressive damage of the macula, the center of retina responsive for central and precise vision.Genetic risk factors response to the environment stimulants, such as oxidative stress resulting in drusen formation, inflammation and abnormal vascular growth. The unique nature of the eye leads to an abnormal burded of both degraded lipid products and oxidative stress, leading to relatively greater burden of oxidized lipid biproducts such as oxPLs. There are two forms of advanced AMD: graphic atrophy (GA) (upper right) and choroidal neovascularization (CNV) (lower right).
Despite the prevalence of this disease, its etiology remains largely unknown. A growing amount of evidence has indicated that the pathogenesis and progression of AMD result from a combination of genetic risk factors and environmental insults such as smoking, UV exposure and microbial assault [6]. However, as with much of the underlying physiology of AMD, the interplay between environmental factors linked to AMD and genetic variants resulting from risk-associated genetic variation remains a mystery. Efforts to understand the pathological interplay of multifactorial systems have led our lab to explore the specific interactions of risk-associated genotypes associated with complement factor H (CFH) proteins and consequences of oxidative stress, specifically those related to lipids. We found that the CFH genetic variation CFHY402H, which is association with increased AMD risk, demonstrated decreased interactivity with oxidation-modified lipids. As the association of CFH with these molecules restricted their innate inflammatory capacity, this diminished affinity disrupts of the tightly controlled inflammatory homeostasis of the eye, leading to increased inflammatory burden which is central to AMD pathogenesis [8]. As the accumulation of chronic, low-level inflammation is exacerbated over time by the accumulation of oxidation products, the result is a gradual progression of disease pathology over the course of life, eventually causing tissue damage that permanently impairs the central vision, leading to blindness. Studies such as these which uncover a novel interplay between genetic and environmental disease contributors provide insight into the complex pathology involved in AMD, and are critical to furthering our understanding towards creating the next generation of approaches to AMD therapeutics. The following review investigates further the specific actors within genetic variations and environmental stressors that lead to the irreversible vision loss caused by AMD.
Genome Wide Association Studies (GWAS) have identified several genes and/or single nucleotide polymorphisms (SNPs) that associate with AMD (Figure 3). However, as mechanistic and genetic investigations of the underlying triggers of AMD pathology continue to evolve, the polygenicity and complex interplay with environmental triggers increasingly portrays AMD as a disease where variable combinations of factors converge on the same pathogenic road towards macular degeneration and vision loss. Understanding these convergence points, and the commonalities of these variable disease effectors, remains the scientific mystery likely to hold the key to understanding the true underlying mechanisms of this disease. These understanding will undoubtedly be central to developing the next generation of truly efficacious interventions. This section will provide an overview of the more substantive identified genetic contributors to AMD, as well as a brief discussion of the putative mechanistic contributions these genetic factors afford to AMD. This will be followed by a discussion of environmental contributions, with a focus on the most highly validated environmental contributor to AMD, oxidative stress. Finally, this review will discuss the potential interplay between these factors, providing additional mechanistic insight into both aspects of AMD risk. Table 1 shows the genes and SNPs that associate with AMD risk. Here we also show a Manhattan plot, which display genetic variants in genes and chromosome regions associated with AMD.
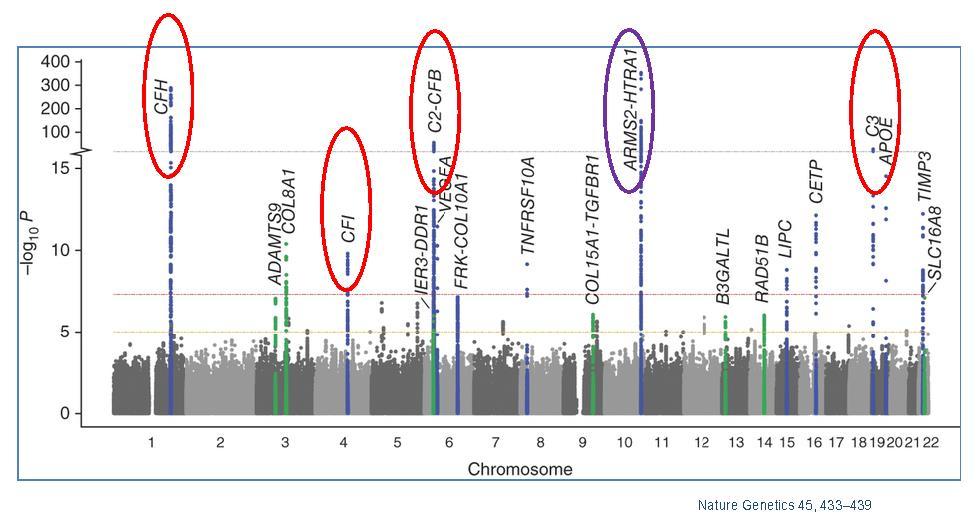 Figure 3. Manhattan plot showing the summary of genome-wide association results in the discovery GWAS sample. The significance of association for genetic variants including single nucleotide polymorphisms (SNPs) in a genome-wide association analysis is indicated by the P values in log scale. The data set are plotted for SNPs on each chromosome with P < 5 × 10−8 labeled with the gene. Red circles indicate genes in complement pathway; purple circle indicates the HTRA1/ARMS2 loci. Adapted from [9].
Figure 3. Manhattan plot showing the summary of genome-wide association results in the discovery GWAS sample. The significance of association for genetic variants including single nucleotide polymorphisms (SNPs) in a genome-wide association analysis is indicated by the P values in log scale. The data set are plotted for SNPs on each chromosome with P < 5 × 10−8 labeled with the gene. Red circles indicate genes in complement pathway; purple circle indicates the HTRA1/ARMS2 loci. Adapted from [9].
| Genes | Variants (SNP) | Full Name | Function | Position | Odds Ratios | References |
| ABCA4 | rs1800553, rs1800555 | ATP-binding cassette, sub-family A (ABC1), member 4 | Photoreceptor specific expression; transport of N-retinylidene-PE across the outer segment disc membrane | 1p22 | N.A. | [10,11,12] |
| APOE | rs429358, rs7412 | apolipoprotein E | lipid and cholesterol transport | 19q13 | ε2 ORhomo = 1.046; ε4 ORhomo = 0.847 (Thakkinstian); ε4 ORhomo = 0.35-0.53, 0.847 (separate report-Francis) | [13,14] |
| ARMS2 /LOC387715* | rs10490924, in/del (372_815delins54) | age-related maculopathy susceptibility 2 | no known function; protein localized to mitochondrial outer membrane | 10q26 | ORhomo = 8.59 (rs10490924) | [12,15,16,17] |
| HTRA1* | rs11200638 | human High Temperature Requirement A1 | trypsin-like serine protease | 10q26 | ORhomo = 6.92, 7.46 | [6,18,19] |
| C2/CFB | rs9332739 (c2), rs4151667(CFB), rs641153 (CFB) | complement 2/ complement factor B | regulation of complement activation | 6p21 | ORhetero = 0.21-0.45 | [6,20] |
| C3 | rs2230199, rs1047286 | complement 3 | Innate immunity (alternative complement pathway activator, classical pathway component) | 19p13 | ORhomo = 1.93-3.26 | [21,22,23] |
| CETP | rs1864163 | Cholesteryl ester transfer protein | Transfer cholesteryl esters between lipoproteins | 16q21 | ORhomo = 1.2 | [24] |
| CFH* | rs1061170 | complement factor H | inhibitor of alternative complement pathway | 1q32 | ORhomo = 6.35 | [25,26,27,28] |
| CFHR1 /CFHR3 | 84K bp deletion | complement factor H-related 1, 3 | exact function unknown, possible overlapping function with CFH | 1q31-q32 | ORhomo = 0.29 | [29,30] |
| CFI | rs4698775 | Complement factor I | regulation of complement activation | 4q25 | ORhomo = 1.1 | [31] |
| CX3CR1 | rs3732378 | chemokine (C-X3-C motif) receptor 1 | Inflammatory (chemokine receptor) | 3p21 | ORhomo = 1.98-2.70 | [32,33] |
| ERCC6 | rs3793784 | excision-repain cross-complementing, group6 | DNA repair | 10q11 | ORhomo = 1.6 | [34] |
| LIPC | rs920915 | Hepatic lipase | Lipid metabolism | 15q21-23 | ORhetero = 1.1 | [24,35/a>,36] |
| TIMP3 | rs9621532 | Tissue inhibitor of metalloproteinase | Complexes with inactive metalloproteinase | 22q12 | ORhomo = 1.31-1.91 | [37,38,39] |
| TLR3 | rs3775291 | toll-like receptor 3 | Innate immunity (encodes protein to recognize viral dsRNA) | 4q35 | ORhomo = 0.44-0.61 | [40,41] |
| TLR4 | rs4986790 | toll-like receptor 4 | Innate immunity (encodes bacterial endotoxin receptor) | 9q32-q33 | ORhetero = 2.65 | [32,42,43] |
| VEGFA | rs833069, rs1413711 | vascular endothelial growth factor A | Angiogenic and vasculogenic growth factor | 6p12 | ORhomo = 5.29 (rs833069), ORhomo = 2.40 (rs1413711) | [44,45,46] |
| *ORhomo indicates odds ratio from homozygous; ORhetero indicates odds ratio of heterozygous. | ||||||
 DownLoad: CSV
DownLoad: CSVThe best characterized region of genetic risk associated with AMD has been repeatedly demonstrated to localize within a specific loci on chromosome one. Within this region, genetic variants associated with the alternative complement pathway in multiple forms have been identified. For example, the risk genotype of rs1061170 that causes amino acid changes in CFH 402Y to H (CFHY402H) increases AMD risk between 2 to 4-fold for heterozygote carriers (only one allele of chromosome carries the risk variant) and 3 to 7-fold for homozygotes (both chromosome alleles carry the risk variant) [26,27]. As part of the innate defense system, the complement system is tightly regulated by many of its component factors, such as CFH and CFH related proteins (CFHRs) in modulating the alternative complement pathway. CFH, which consists of 20 short consensus repeats (SCRs), preferentially binds to host cells through glycosaminoglycans to protect host tissue from complement-mediated damage. CFHRs are composed of variation combinations of the SCRs, but all lack SCR1-4 (Figure 4 from [47]). Nevertheless, as the homology may suggest, CFH and CFHRs play similar, yet intricate, roles in regulating the complement pathway [48,49]. In the immune privileged eye, under the condition of non-infectious settings, oxidatively modified materials can be recognized “non-self” and removed by the complement system to maintain homeostasis [50,51]. Similar to naturally occurring antibodies, CFH plays an anti-inflammatory role in the eye by restricting immune activation in response to these molecules [8]. The risk-associated CFHY402H mutation reduces the affinity of CFH to bind such molecules, thereby reducing its ability to maintain immune homeostasis in the eye.
 Figure 4. The genetic (A) and domain structures (B) of human CFH and CFHRs (adapted from [47]).
Figure 4. The genetic (A) and domain structures (B) of human CFH and CFHRs (adapted from [47]).
While wild-type CFH has demonstrated protective benefits in AMD, expression of CFHR1 and CFHR3 demonstrate a surprising and opposite effect in their ability to increase risk of AMD occurrence [52,53]. Paradoxically, loss-of-function mutation or deletion of complement-protective CFHR1/3 (protective for AMD) is actually pathogenic in other conditions such as C3 glomerulopathy [54] and hemolytic uremic syndrome (HSU) [55]. In the latter case, homozygous deletion of CFHR1/CFHR3 is strongly associated with the development of factor H auto-antibodies (DEAP HUS) [49,55,56,57]. The identification of CFHR1, but not CFH, in AMD drusen and Bruch’s membrane indicates an important distinction in their ability to contribute to lipid accumulation in RPE, which is likely to contribute to the inability of CFHR1/3 to maintain inflammatory homeostasis in the eye. The opposing contributions of CFH and CFHRs in conferring disease risk indicate that the suppression of complement activation by CFHRs may be mechanistically uncoupled, at least in part, from their role in promoting AMD risk (Figure 5). However, other polymorphisms in non-coding regions of CFH, or in nearby genes encoding other complement factors, also demonstrate comparably strong association with disease susceptibility. For example, two genes within the major histocompatibility complex class III region have been identified as AMD risk variants; factor B (CFB) and complement component 2 (C2) [20,25]. As genetic variations in this system can both promote and protect against disease risk, it appears that any disruption of this system can have a prominent effect on disease. In general the genetic variations in the complement pathway that contribute to disease risk putatively fall into 2 categories of disrupted control of complement-mediated immune activity: (i) alterations that lead to unchecked hyperactivation of the complement pathway, (ii) alterations that restrict the checks on protection of endogenous molecules from unwanted complement attack. In each case, either loss of “self” protection or undesirable complement activity contribute to chronic increases in low-level inflammation that can then contribute to AMD progression over time. However, the specific functions of these mutations in the specific context of environmental modifications of lipids is of particular interest. This will be discussed further in discussion of oxidative stress contributions to AMD risk.
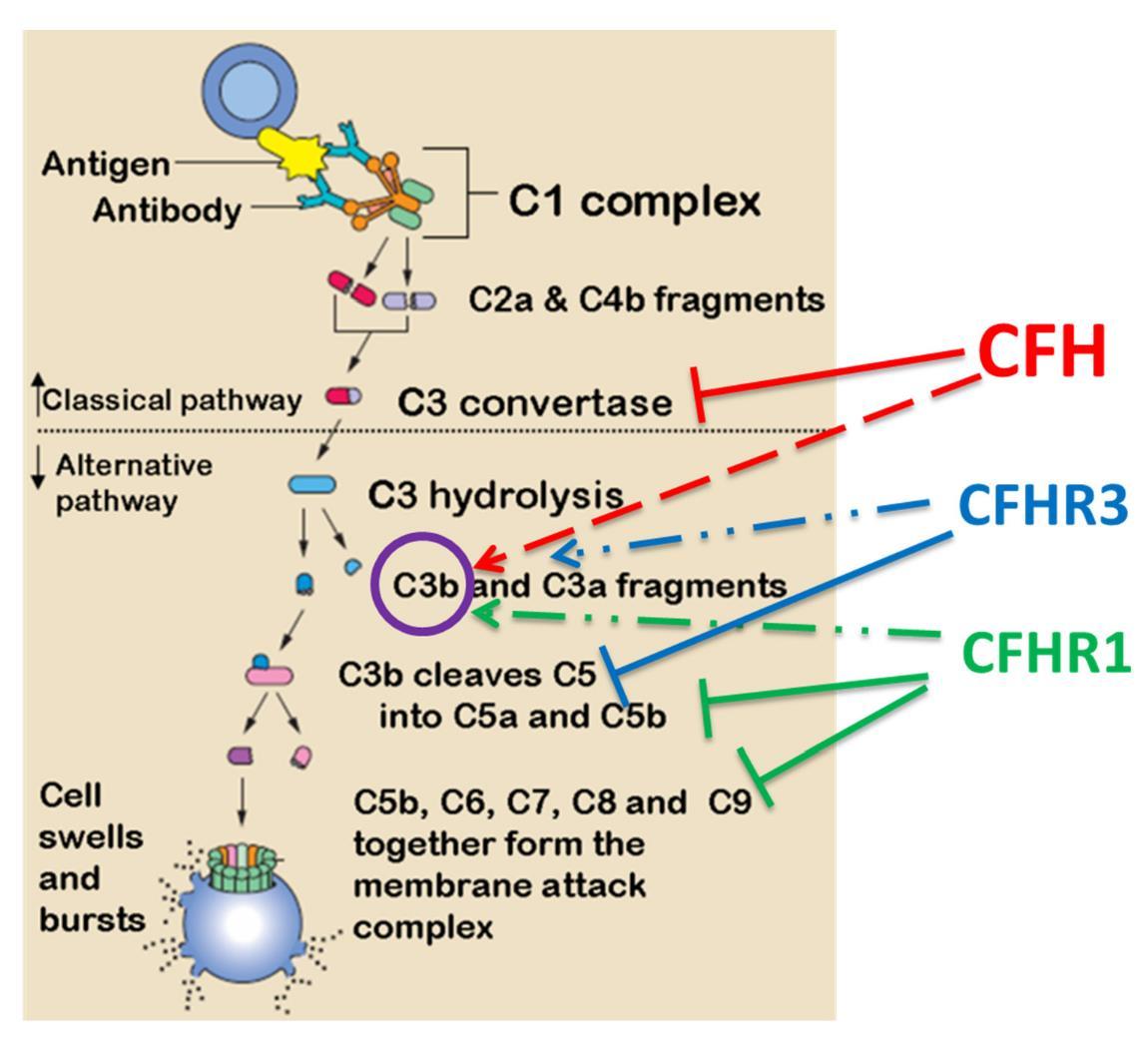 Figure 5. Putative role of CFH and CFHRs in regulating classical or alternative complement pathways. CFH interacts with C3 convertase and also with C3b. CFHR1/3 inhibit later stages of the cascade and their deficiency of would results in a loss of complement control but enhances local regulation CFH.
Figure 5. Putative role of CFH and CFHRs in regulating classical or alternative complement pathways. CFH interacts with C3 convertase and also with C3b. CFHR1/3 inhibit later stages of the cascade and their deficiency of would results in a loss of complement control but enhances local regulation CFH.
In addition to variations associated with the complement system, genes involved in lipid metabolism arise as the next common system associated genetic variants with links to AMD risk [58,59]. As disruption of lipid homeostasis in the eye in terms of accumulation, improper degradation, aggregated storage, etc., are hallmarks of AMD pathology, this is perhaps unsurprising. The accumulation of lipids, particularly in terms of the formation of drusen, leads to chronic inflammation in the eye, further converging the handling of lipids into the “usual suspects” of AMD progression. The cholesterol transport protein apolipoprotein E is well-established as perhaps the strongest genetic factor related to the most well-known age-related pathologies of our time; atherosclerosis and Alzheimer’s Disease (AD) [60,61,62]. In humans, the ApoE gene is represented by 3 alleles; ApoE2, ApoE3, and ApoE4. In AD, ApoE4 is of the strongest genetic factors associated with disease risk, while ApoE2 is protective. Surprisingly, in AMD the opposite is true with ApoE2 being associated with increased risk of AMD and ApoE4 serving a protective role [63]. All 3 ApoE variants can be found in the RPE and Bruch’s membrane, but while ApoE4 is protective and correlates to decreased inflammation and macrophage recruitment, ApoE2 results in increased macrophage infiltration and inflammation [64]. While the complexity of this paradox is likely multifaceted, the affinity for different ApoE isoforms for amyloid-β oligomers is widely believed to play at least some role in the susceptibility to AD. As such, the opposite effect on disease risk in AMD is likely due to similar distinctions in affinity for molecules in the eye that cause inflammation. This will be discussed further in later sections.
While ApoE proteins are well established contributors to AMD, they are not the only lipid-handling proteins implicated as contributors of genetic risk to AMD. Cholesterol ester transfer protein (CETP), lipoprotein lipase (LPL) and hepatic lipase (LIPC) have all recently been reproducibly implicated as factoring into AMD risk [9,35,36] These proteins are central to proper handling and degradation of lipoproteins, further demonstrating the vital nature of lipid homeostasis in AMD prevention. Similarly, polymorphisms in the cholesterol exporter ABCA1 also demonstrates increased risk for AMD [36]. As lipid homeostasis increasingly demonstrates a central theme in AMD pathogenesis, we will likely continue to see additional lipid-related genes with AMD influence. However, as the number of genes continues to grow, it is unlikely that any single gene will provide value as a therapeutic target. Rather, promotion of lipid homeostasis/proper lipid handling will likely prove a more rational approach to the disease.
Another prominent genetic association with AMD are SNPs in or near the promoter region of the high temperature required factor A1 (HTRA1) or age-related maculopathy susceptibility protein 2 (ARMS2) genes, including rs11200638 rs10490924, rs11200638 and rs2293870 on chromosome 10, which are perhaps the most well-documented genetic associate with neovascular AMD [15,18,19]. Within this genetic region lies a common disease haplotype, TAT-tagged by rs10490924, rs11200638 and rs2293870, which encompasses HTRA1/ARMS2 and is significantly associated with the risk for AMD [15]. HTRA1 is a serine protease that has been shown weaken extracellular matrix (ECM) [65]. In addition, our previous study has characterized its ability to function as a transcriptional enhancer of VEGF expression, which has obvious implications for neovascularization. In the Bruch’s membrane, HTRA1 enzymatic activity compromises the compartmental integrity of the basal retina, which can leave the tissue susceptible to the aberrant vascularization from increased levels of VEGF. Additionally, the abnormal levels of HTRA1 found in AMD patients that causes Bruch’s membrane deterioration also leads to stimulation of inflammation of the RPE, which has also been observed in vitro [65].
Another protease contributor to AMD is the tissue inhibitor of metalloproteinases 3 (TIMP 3), which belongs a group of peptidases involved in degradation of the ECM. Various forms of stimulation such as oxidative stress and inflammatory cytokines can induce TIMP3 expression, and genetic variants in this gene have been associated with AMD [37,38,39] and the highly AMD-related autosomal dominant disorder Sorsby’s fundus dystrophy (SFD) [66]. However, the role of TIMP3 in AMD is somewhat controversial as some have proposed, due to the penetrance of TIMP3 in subpopulations of patients with macular degeneration, that this particular gene may represent a specific and distinct disease subset [67].
In summary, the genetic variants at the CFH and HTRA1/ARMS2 loci contribute to major genetic risk for AMD, which lead to growing functional study aiming to elucidate the molecular mechanisms underlying AMD pathogenesis, in particular how such changes interplay with the environmental risk factors. As the number of genetic studies investigating AMD continues to grow, evidence continues to emerge indicating additional rare coding variants also involved in disease, allowing us to better pinpoint causal genes within known genetic loci. As such results are experimentally confirmed, we increasingly see the need for large sample sizes to detect new loci and genes that can inform on disease pathology [68].
AMD risk has both environmental and genetic contributions. However, as an age-related disease, it also shares common risk contributors with other chronic health ailments related more to lifestyle or medical comorbidities as opposed to environmental exposure. Conditions such as obesity, sendentary lifestyle, high cholesterol and high blood pressure have well-defined influence on promoting AMD risk [69,70,71,72,73,74,75,76]. However, the ability of such factors to promote AMD progression is highly non-specific as these factors influence a variety of chronic, age associated pathologies such as atherosclerosis, diabetes, AD, etc. As such, for the sake of this review we will focus primarily on the more fundamental environmental and biological contributors even as some may be secondary to these more broadly encompassing lifestyle contributions.
When considering the global list of correlated factors contributing to AMD, it becomes apparent that many of these factors (i.e.: sunlight exposure, diet, smoking, vitamin D levels, etc. [75,77,78]) have well-documented effects on oxidative stress and its consequent inflammation. Additionally, the downstream effects of oxidative stress have a variety of disparate disease-related consequences that can influence disease progression through several avenues. As such, the following will focus to some extent on the general concept of oxidative stress and its role in other contributing aspects of AMD progression to highlight the centrality of this factor in this disease.
The contribution of oxidative stress to age-associated pathology is a common trend in many diseases such as cancer, atherosclerosis, Alzheimer’s disease and Parkinson’s disease [79,80,81,82,83], and is a natural consequence of many of the lifestyle-associated risk factors discussed above. The centrality of oxidative stress as a disease contributor to AMD was highlighted by the Age-Related Eye Disease Study (AREDS); a major clinical trial sponsored by the National Eye Institute which was designed to learn more about the natural history and risk factors of AMD. The study specifically evaluated the effect of high doses of vitamin C, vitamin E, beta-carotene and zinc on the progression of AMD and concluded that a variety of antioxidant agents ameliorate AMD. This study was the first to confirm in humans via rigorous assessment the importance of oxidative stress in initiation and progression of the disease ?[69,76]. This perspective has been supported anecdotally by several studies pinpointing smoking is the top environmental risk for developing AMD in all age and ethnic groups [74,84,85,86,87,88,89], as well as in other ocular diseases [90].
Oxidative stress/damage in the eye can be occurred in many forms via a variety of stimuli other than smoking [91]. In human eyes, local exposure sunlight combined with the high local oxygen content, which is higher than other tissues, leads to a high predisposition for oxidative burden. When combined with the systemic exposure to oxidative stressors incurred via lifestyle choices or other contributors, the relative burden of oxidative stress can rapidly become disproportionately high. While oxidative stress has been linked to a variety of specific and general modes of inflammatory promotion, oxidative stress has specific mechanistic consequences in the eye directly related to AMD pathogenesis ?[81,82,91,92,93].
In the eye, retinal photoreceptor outer segments are constantly turned-over by retinal pigment epithelium (RPE) and enriched with polyunsaturated fatty acid (PUFA) phospholipids such as phosphatidylcholine (PC), which results in a constant supply of lipid products to be cleared and handled by the retina [94,95]. While the system is capable of maintaining such turnover, the relatively high level of oxidative burden in the eye locally and systemically results in a ready pool of lipids available for oxidative modifications. This combination can give rise to a variety of lipid modifications, particularly oxidized phospholipids (oxPLs) arising from turned over PUFAs. For example, oxidative modification of a phospholipid such as PC results in a conformational change of the head group exemplified by oxidation of 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine (PAPC) into 1-palmitoyl-2-(5'-oxo-valeroyl)-sn-glycero-3-phosphocholine (POVPC) (Figure 6). This, and similar, modifications can change otherwise ubiquitous and benign biomembrane molecule into neo-epitopes that begin to resemble that of bacterial membrane proteins, which can incite undesirable attention from immune cells [8,96].
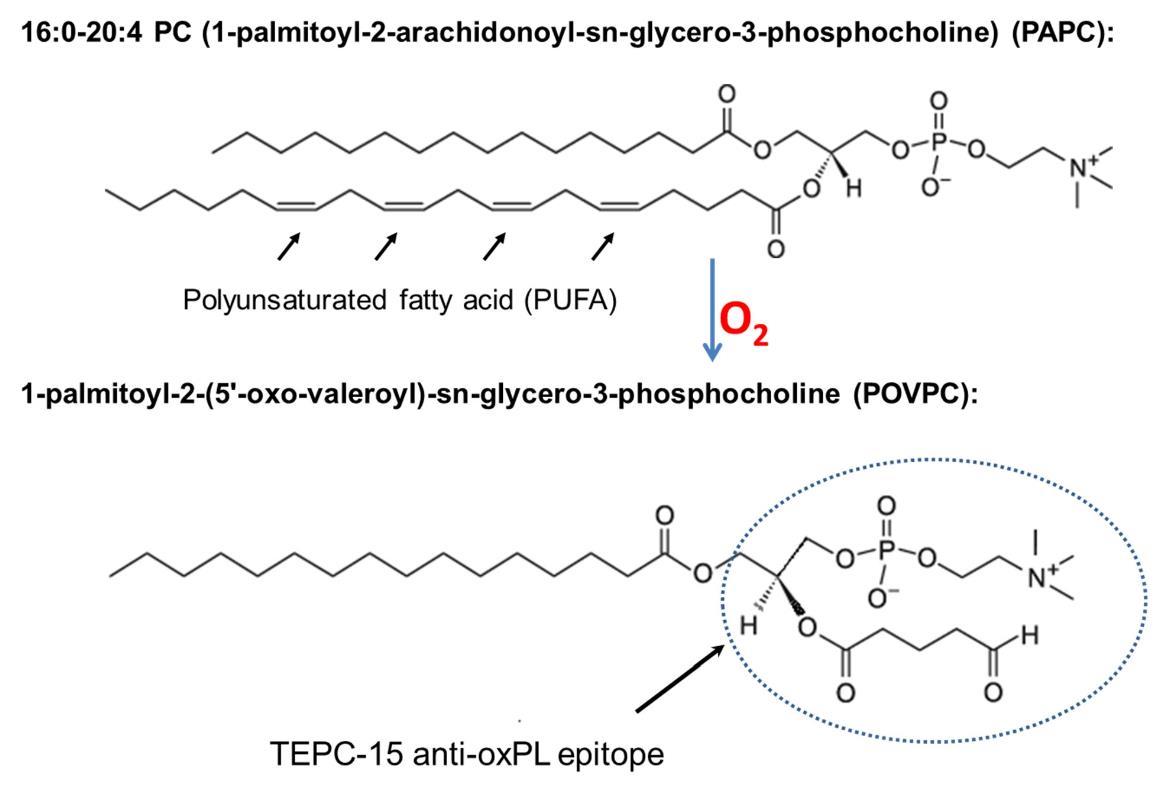 Figure 6. Chemical structure of PAPC, a common membrane phospholipid in the retina, and conversion to oxidatively modified POVPC, which recognized by a monoclonal antibody specific to oxPLs.
Figure 6. Chemical structure of PAPC, a common membrane phospholipid in the retina, and conversion to oxidatively modified POVPC, which recognized by a monoclonal antibody specific to oxPLs.
Oxidation of lipoproteins also produces oxysterols such as 7-ketocholesterol, 25-hydroxycholesterol, etc. When generated in vivo, similar to oxPLs, these are pro-inflammatory and have been shown to contribute to AD through their ability to alter cholesterol homeostasis in the brain [97], as well as atherosclerosis via activation of macrophages and stimulation of foam cell formation [98]. Many oxysterols in the eye have been shown to be pro-inflammatory and cytotoxic to both photoreceptors and RPE cells [99].
Several in vitro and in vivo studies have established the association of AMD with a variety of such peroxidation-degraded lipids. During lipid peroxidation, the reactive oxygen species attack polyunsaturated fatty acids causing the breakdown of double bond that results in a variety of degraded, oxidized lipid byproducts [100]. For example, enhanced lipid peroxidation necessitated in the retina for proper lipid metabolism creates several breakdown products such as malondialdehyde (MDA), malondialdehyde-acetaldehyde (MAA) [101] and POVPC. These newly modified molecules are very reactive and can facilitate interaction with a variety of matrix and structural proteins, as well as cellular membranes, to form adducts. These adducts form molecular moieties which are inherently antigenic, which can lead to improper targeting of healthy cells/molecules by the immune system causing inflammation. Consequently, studies have identified adducts of MDA [101], oxidized phosphocholine (oxPC), carboxymethyllysine (CML) [102], pentosidine [103], and carboxyethylpyrrole (CEP) [104] in AMD drusen. The fact that these oxidatively-induced modifications of autologous proteins convert ubiquitous molecules into antigenic, inflammatory stimulants is central to understanding the functional mechanism of these modifications as well indicative of the potent role of oxidative stress in AMD pathology.
In addition to its ability to modify lipid structure, oxidative stress can also lead to DNA damage, particular within mitochondria where high ROS are potent agents to mitochondrial DNA [105]. Interestingly, it has been shown that DNA or RPE cells in AMD patients exhibit extensive DNA damage which leads to an inflammatory response [106]. This DNA damage-related inflammation has been previously shown to not only associate with AMD, but also with aging in general [107,108]. As a result, protection of mitochondrial DNA from oxidative and other forms of damage has been proposed as a novel therapeutic strategy to slowing the progression of AMD [109].
In sum, in AMD the contribution of oxidative stress combined alterations in lipid handling caused by with the known link to genetic changes in lipid handling and inflammatory modulation indicates that inflammatory homeostasis in the eye via tight regulation of oxidized lipid products, and non-lipid byproducts, may be central to this disease.
It is now clear that oxidative modifications of molecules within the eye such as those discussed above are pro-inflammatory and promote the progression of early AMD drusen to CNV or GA. The question then arises, what are the mechanisms that translate these inflammatory signals in AMD pathology? It is known that generic retinal inflammation is characteristic of a number of pathologies of the eye, and while such inflammation is undoubtedly central to AMD, chronic eye inflammation does not always lead to AMD [110]. As such, identifying specific triggers and mechanisms of inflammation, as well as the participating cell types, is foundational to our understanding of this disease. We have previously discussed the prominent role of oxPLs in AMD due to the promotion of inflammation. The mechanism by which this occurs is a results of the ability of such molecules to recruit T-cells and monocytes to the subretinal tissue, where monocytes then differentiate into macrophages. These macrophages take on a strong pro-inflammatory phenotype, even sometimes resembling the morphology of foam cells readily found in atherosclerotic lesions, leading to a potent inflammatory burden within the retina [8,111]. While not directly proven to influence the transformation of early AMD to CNV, oxidized lipids have also been reported to directly affect growth, differentiation, and survival of vascular cells, which may be why some patients with wet AMD fail to respond to anti-VEGF therapy [112]. In fact, studies indicate that even early forms of oxidized lipoproteins (e.g. minimally modified LDL) cause changes in gene expression (e.g. activating NFkB-like factors) of vascular cells, leading to the initiation and maintenance of an inflammatory response that could contribute to conversion of early drusen into advanced CNV [113,114].
Despite the overwhelming evidence that complement pathway gene polymorphisms strongly associate with AMD disease risk, the mechanism and pathways through which oxidative stress affect AMD are not yet clear. Our lab has recently identified a novel mechanistic interplay between the genetic variations in CFH associated with disease risk and oxidative modifications of lipids. Although eye is an immune privileged organ, CFH protein have been observed in retina and AMD drusen (Figure 7) indicating its role that may not directly involve in regulation complement activation. Wild-type CFH demonstrates a substantially increased affinity for oxidatively modified lipids such as oxPLs and malondialdehyde (MDA) compared to native, un-oxidized lipids [8,115,116]. As these oxidized lipids have inflammatory capacity, the interaction of CFH acts to restrict this inflammatory potential in a similar fashion to naturally occurring antibodies of the innate immune system [115]. Risk-associated genetic variants either decrease the affinity of CFH for these oxidation products, or displace CFH with CFH- related proteins that lack the same anti-inflammatory capacity. As such, the oxPLs burden has a higher propensity for RPE interaction leading to inflammation and drusen formation in these patients.
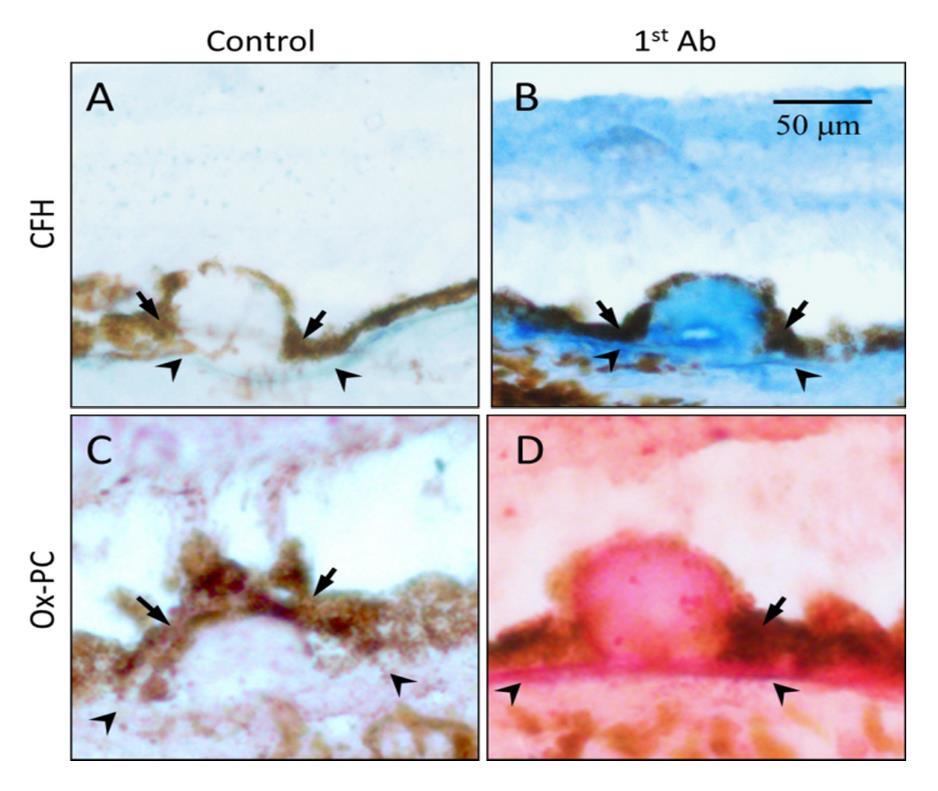 Figure 7. Co-localization of oxPC with CFH in human AMD lesions. Immunohistochemistry of serial sections of an AMD eye stained with antibodies to CFH (Panel B, blue color) or oxPC (Panel D, pink color). Omission of 1st antibody served as a negative control (Panels A and C). Size bars: 50 microns.[8].
Figure 7. Co-localization of oxPC with CFH in human AMD lesions. Immunohistochemistry of serial sections of an AMD eye stained with antibodies to CFH (Panel B, blue color) or oxPC (Panel D, pink color). Omission of 1st antibody served as a negative control (Panels A and C). Size bars: 50 microns.[8].
Innate immune system protect host from lethal pathogenic microbial assults before adaptive innunity kicks in by recognizing pathogen-associated molecular patterns (PAMPs). Previously, studies have made an interesting finding that in addition to defending the host from microbial infection, natural antibodies, which are a branch of innate immunity, can shield host from oxidative damages, thereby preventing un-wanted inflammation, particularly in response to oxidation-specific neo-epitopes [96]. These antibodies prevent the immune system from inappropriately reacting to self-proteins as pathogens, which is normally an interaction restricted to PAMPs on microbial surface or debris. Oxidized lipid products can sometimes mimic PAMPs, requiring intrinsic mechanisms to control for attack of these non-pathogenic structures. For example, mouse natural antibody TEPC-15 (aka T15), which protects neonatal animals from fetal pneumococcal infection before adaptive immune response, recognizes and interacts with phosphocholine (PC) n-linked on the cell wall of certain bacteria such as S. pneumococci. However, this anti-PC antibody does not recognize the PC structure o-linked to mammalian cell membrane phospholipids. After oxidative modification of phospholipids as illustrated in the Figure 6, the conformational change will present modified PC head-group to the host immune system as a neo-antigen resembling those on bacterial cell walls. Therefore, these modified cellular and molecular structures from endogenous oxidative damage are also called damage associated molecular patterns (DAMPs). Interestingly, after oxidative modification, when phospholipids on cell membrane become oxPLs, the same PC epitope can be recognized and interacted by T15. As such, T15 is able to interact with endogenous oxPC on cell membranes or lipoproteins and maintain homeostasis by turnover of unhealthy cells and macromolecules such as to remove apoptotic cells and oxLDL [96,115].
Even though the eye is an immune privileged organ, the presence of resident immunocompetent cells (e.g. microglia), and humoral factors such as cytokines, adhesion molecules, auto-antibodies, and acute phase proteins lead to a somewhat self-contained immune system that can play important roles in determining the inflammatory status of the retina. However, understanding the specific cellular contributors and signals unique to AMD is important in the understanding of this disease. Unsurprisingly, a panel of pro-inflammatory cytokines and chemokines, including IL-1, IL-6, IL-8, TNF, INF-γ, MCP-1, have been shown to accelerate the AMD progression [117]. In addition, tissue factors that have pro-angiogenic activity, such as VEGF A-E, platelet-derived growth factor (PDGF), placental growth factor (PlGF), hepatocyte growth factor (HGF), and fibroblast growth factor-2 (FGF-2), are also involved in CNV formation [118].
Recently it was shown that CFHY402H patients expressed significantly greater levels of INFγ-inducible protein-10 (IP-10) and eotaxin, perhaps indicating such factors may be useful as vitreal biomarkers of early AMD [119]. As this was uniquely identified in this specific genetic background, future studies to assess the incidence of elevation of these markers in non CFHY402H AMD patients will be of great interest. Similarly, it has also been shown in CFHY402H patients that macrophage infiltration is likely stimulated by elevated levels of vitreal granulocyte macrophage colony-stimulating factor (GM-CSF), leading to detection of choroidal macrophages in the postmortem human eye [120]. The role of these macrophages is yet unclear, as these cells can accumulate in sites of damage or disease without contributing to disease progression, in some cases even serving a beneficial purpose. However, a recent study demonstrated a large population of CD163 positive cells in the retina of wet and dry AMD postmortem human samples [121]. As CD163 is associated with potently inflammatory immune populations, it is likely that macrophages present in the retina of AMD patients are not serving any putative disease restricting purpose. Regardless, these specific, localized inflammatory events continue to provide insight into the unique players involved in promotion of inflammatory disease within an immune-privileged tissue. As such, further characterization and mechanistic elucidation could identify novel therapeutic avenues by which disruption of disease-associated cellular recruitment, or inflammatory status of infiltrating cellular populations, could slow the progression of vision loss.
The infiltration of macrophages from the peripheral circulation is a unique and interesting component of AMD pathology. The role of these macrophages, as well as the specific functions of the pro-inflammatory (M1) and inflammation suppressing (M2) phenotypes in regulating AMD pathology, is currently an important topic of active investigation [11]. M1 macrophages in the retina contribute to the elevated levels of inflammatory cytokine/chemokines known to be associated with AMD such as IL1, IL6, IL8, and the production of VEGF. Conversely, while M2 macrophages do not express the inflammatory milieu characteristic of M1, they do produce pro-angiogenic factors, such as basic fibroblast growth factor (FGF-2), insulin-like growth factor-1 (IGF-1), and placental growth factor (PGF) [10]. As a result, while M2 macrophages are largely considered beneficial to disease, this pro-angiogenic function could still contribute to wet AMD progression despite expression of anti-inflammatory cytokines such as IL-10 and IL-18 that have shown to have protective effect in regard to AMD risk [122].Additionally, while M1 macrophages are commonly considered pathogenic, their expression of the cytokine IL-18 has been shown to combat theneovascular effects of VEGF via attenuation of CNV in rodent models, as well as in nonhuman primate models of wet AMD, thereby indicating a potential therapeutic role for M1 macrophages in this pathology [123,124]. Future work should elaborate on the intricacy of macrophage function in these diseases, and the specific contributions of M1 vs. M2 phenotypes.
The commonalities between wet and dry AMD are numerous in their early stages. The strong association of complement pathway genes with AMD susceptibility [125] and the presence of complement proteins in drusen indicate that AMD is likely, at least in part, a chronic inflammatory disease involving abnormal regulation of complement and immune system [25,126,127,128,129,130,131]. Proteomic studies have found that many oxidatively modified proteins and lipids as well as immunoglobulins in AMD drusen [91,131,132]. Such oxidative materials including oxPLs are strong stimulants to the immune system resulting in promoting inflammation and activating the complement cascade in RPE cells. Thus in early and intermediate stages of AMD, the immune responses to oxidative damage play important roles in RPE apoptosis and lipids accumulation leading to drusen formation.
Despite the early hallmark pathology for both forms of wet and dry AMD to be commonly held, a divergence point seems defined by the degradation of the Bruch’s membrane and parallel development of malformed vasculature infiltration. The clearest differentiating contributor to this pathological distinction is the role of VEGF. However, VEGF levels are most likely a secondary consequence of the pathological mechanisms at play as opposed to directly related to disease progression due to some form of unwanted, and untriggered, overexpression. The SNPs in or near the promoter region of the high temperature required factor A1 (HTRA1) or age-related maculopathy susceptibility protein 2 (ARMS2) genes on chromosome 10 include rs11200638 rs10490924, rs11200638 and rs2293870 are the strongest genetic contributor to wet AMD and these result in increased production of VEGF in the retina [19,65]. In addition to its characterized ability to function as a transcriptional enhancer of VEGF expression, which has obvious implications for CNV, HTRA1 is also a serine protease that has been shown weaken ECM [65,133]. In the Bruch’s membrane, this enzymatic activity compromises the compartmental integrity of the basal retina, which can leave the tissue susceptible to the aberrant vascularization that results from increased levels of VEGF. Additionally, the abnormal levels of HTRA1 found in AMD patients that causes Bruch’s membrane deterioration also leads to stimulation of inflammation of the RPE, which has also been observed in vitro [65]. While it has not been tested, TIMP3 may also contribute to CNV pathogenesis in a similar fashion.
Although the specific role of the macrophage lineage cells at different stages of AMD is still controversial, these components of innate immune system play an important role in neovascular AMD [134,135]. The macrophages recruited in the retina are from two sources. The first one belongs to microglia, which are bone marrow-derived resident macrophages recruited to neural tissue during retinal development. They provide immune surveillance in the inner retina and have been associated with AMD [136,137]. The second group is from circulating monocytes that can be recruited from the blood vessels to sites of inflammation by specific chemokines and cytokines, which are stimulated by oxidative in the retina and RPE [138,139]. Regardless of the source, macrophages can undergo further differentiation depending under the oxidative microenvironment and eventually perform their effector functions [140,141]. For example the presence ofextracellular nitric oxide synthase (iNOS) is associated with macrophage recruitment to Bruck’s membrane and alteration in the immunophenotype of resident choroidal macrophages [135,142]. In the presence of interferon-gamma (IFN-γ), macrophagesare activated as proinflammatory M1 macrophages, which produce tumor necrosis factor alpha (TNF-α) and interleukin-12 (IL-12) and are associated with tissue damage [143].
In the past decade, there has been an unprecedented increase in the understanding of genetic and environmental contributions to AMD. These novel insights have begun to frame a general interplay of extrinsic and intrinsic factors that contribute to important homeostatic aspects of the eye such as inflammation and lipid processing, but the precise mechanisms contributing to these pathologic disruptions are still poorly understood. By continuing to investigate the functional consequence of genetic variants contributing to AMD such as those found in 1q31-32 and 10q26, which represent genes from the complement system and serine protease family respectively, we can continue to identify biological functions/systems that are repeatedly influenced by identified genetic variants. In turn, we can continue to perform hypothesis-driven research into the potential mechanisms between specific genetic variations and known environmental contributors to disease risk to uncover novel points of intervention within these systems that can preserve vision and improve patient outcomes. While genetic factors are well-characterized as potent contributors to AMD pathogenesis, the disproportionate emphasis on such studies in AMD research is unlikely to catalyze future treatments in an efficient manner. Genetic variations are determined at birth, and while the contribution to risk is quite real, the person-to-person likelihood of developing AMD as a result of carrying these genetic risk factors is often quite minimal, and even then only later in life. As such, these findings are unlikely to provide value proportionate to the effort and funding invested in their acquisition unless subsequently applied to hypothesis-driven studies aimed at elucidating the interplay with environmental or cooperative disease triggers. The in-depth understanding of the interplay between genetic and environmental factors leading to AMD is likely the key to discovering novel therapeutic interventions capable of salvaging vision in these patients.
Peter X. Shaw is supported by the National Eye Institute grants R01-EY-025693 (PXS) and P30EY022589. Hongjun Du is supported by grants from National Natural Science Foundation of China (No. 30973253, 81470654). Xu Xiao is supported by Sichuan Provincial Science and Technology Department supporting fund 2016SZ0054.
The authors declare no conflict of interest, no financial interest to disclose.
| [1] |
C. Alves, F. Corrˆea, T. Ma, Positive solutions for a quasilinear elliptic equation of Kirchhoff type, Comput. Math. Appl., 49 (2005), 85–93. http://dx.doi.org/10.1016/j.camwa.2005.01.008 doi: 10.1016/j.camwa.2005.01.008

|
| [2] |
A. Ambrosetti, P. Rabinowitz, Dual variational methods in critical point theory and applications, J. Funct. Anal., 14 (1973), 349–381. http://dx.doi.org/10.1016/0022-1236(73)90051-7 doi: 10.1016/0022-1236(73)90051-7
|
| [3] |
M. Caponi, P. Pucci, Existence theorems for entire solutions of stationary Kirchhoff fractional p-Laplacian equations, Annali di Matematica, 195 (2016), 2099–2129. http://dx.doi.org/10.1007/s10231-016-0555-x doi: 10.1007/s10231-016-0555-x
|
| [4] |
W. Chen, Critical fractional p-Kirchhoff type problem with a generalized Choquard nonlinearity, J. Math. Phys., 59 (2018), 121502. http://dx.doi.org/10.1063/1.5052669 doi: 10.1063/1.5052669
|
| [5] |
W. Chen, Y. Gui, Multiple solutions for a fractional p-Kirchhoff problem with Hardy nonlinearity, Nonlinear Anal., 188 (2019), 316–338. http://dx.doi.org/10.1016/j.na.2019.06.009 doi: 10.1016/j.na.2019.06.009
|
| [6] |
W. Chen, S. Mosconi, M. Squassina, Nonlocal problems with critical Hardy nonlinearity, J. Funct. Anal., 275 (2018), 3065–3114. http://dx.doi.org/10.1016/j.jfa.2018.02.020 doi: 10.1016/j.jfa.2018.02.020
|
| [7] |
W. Chen, V. Rǎdulescu, B. Zhang, Fractional Choquard-Kirchhoff problems with critical nonlinearity and Hardy potential, Anal. Math. Phys., 11 (2021), 132. http://dx.doi.org/10.1007/s13324-021-00564-7 doi: 10.1007/s13324-021-00564-7
|
| [8] |
I. Ekeland, On the variational principle, J. Math. Anal. Appl., 47 (1974), 324–353. http://dx.doi.org/10.1016/0022-247X(74)90025-0 doi: 10.1016/0022-247X(74)90025-0
|
| [9] |
A. Fiscella, P. Pucci, p-fractional Kirchhoff equations involving critical nonlinearities, Nonlinear Anal.-Real, 35 (2017), 350–378. http://dx.doi.org/10.1016/j.nonrwa.2016.11.004 doi: 10.1016/j.nonrwa.2016.11.004
|
| [10] |
A. Fiscella, P. Pucci, Kirchhoff-Hardy fractional problems with lack of compactness, Adv. Nonlinear Stud., 17 (2017), 429–456. http://dx.doi.org/10.1515/ans-2017-6021 doi: 10.1515/ans-2017-6021
|
| [11] |
M. Khiddi, S. Sbai, Infinitely many solutions for non-local elliptic non-degenerate p-Kirchhoff equations with critical exponent, Complex Var. Elliptic, 65 (2020), 368–380. http://dx.doi.org/10.1080/17476933.2019.1627527 doi: 10.1080/17476933.2019.1627527
|
| [12] | G. Kirchhoff, Mechanik, Leipzig: Teubner, 1883. |
| [13] |
S. Marano, S. Mosconi, Asymptotic for optimizers of the fractional Hardy-Sobolev inequality, Commun. Contemp. Math., 21 (2019), 1850028. http://dx.doi.org/10.1142/S0219199718500281 doi: 10.1142/S0219199718500281
|
| [14] |
S. Mosconi, M. Squassina, Nonlocal problems at nearly critical growth, Nonlinear Anal.-Theor., 136 (2016), 84–101. http://dx.doi.org/10.1016/j.na.2016.02.012 doi: 10.1016/j.na.2016.02.012
|
| [15] |
E. Nezza, G. Palatucci, E. Valdinoci, Hitchhiker guide to the fractional Sobolev spaces, Bull. Sci. Math., 136 (2012), 521–573. http://dx.doi.org/10.1016/j.bulsci.2011.12.004 doi: 10.1016/j.bulsci.2011.12.004
|
| [16] |
N. Nyamoradi, A. Razani, Existence to fractional critical equation with Hardy-Littlewood-Sobolev nonlinearities, Acta Math. Sci., 41 (2021), 1321–1332. http://dx.doi.org/10.1007/s10473-021-0418-4 doi: 10.1007/s10473-021-0418-4
|
| [17] |
P. Pucci, M. Xiang, B. Zhang, Existence results for Schrödinger-Choquard-Kirchhoff equations involving the fractional p-Laplacian, Adv. Calc. Var., 12 (2019), 253–275. http://dx.doi.org/10.1515/acv-2016-0049 doi: 10.1515/acv-2016-0049
|
| [18] |
L. Wang, T. Han, J. Wang, Infinitely many solutions for Schrödinger-Choquard-Kirchhoff equations involving the fractional p-Laplacian, Acta Math. Sin.-English Ser., 37 (2021), 315–332. http://dx.doi.org/10.1007/s10114-021-0125-z doi: 10.1007/s10114-021-0125-z
|
| [19] |
F. Wang, M. Xiang, Multiplicity of solutions for a class of fractional Choquard-Kirchhoff equations involving critical nonlinearity, Anal. Math. Phys., 9 (2019), 1–16. http://dx.doi.org/10.1007/s13324-017-0174-8 doi: 10.1007/s13324-017-0174-8
|
| [20] |
F. Wang, D. Hu, M. Xiang, Combined effects of Choquard and singular nonlinearities in fractional Kirchhoff problems, Adv. Nonlinear Anal., 10 (2021), 636–658. http://dx.doi.org/10.1515/anona-2020-0150 doi: 10.1515/anona-2020-0150
|
| [21] |
M. Xiang, B. Zhang, A critical fractional p-Kirchhoff type problem involving discontinuous nonlinearity, Discrete Cont. Dyn.-S, 12 (2019), 413–433. http://dx.doi.org/10.3934/dcdss.2019027 doi: 10.3934/dcdss.2019027
|
| [22] |
M. Xiang, B. Zhang, X. Zhang, A nonhomogeneous fractional p-Kirchhoff type problem involving critical exponent in RN, Adv. Nonlinear Stud., 17 (2017), 611–640. http://dx.doi.org/10.1515/ans-2016-6002 doi: 10.1515/ans-2016-6002
|
| [23] |
M. Xiang, D. Hu, D. Yang, Least energy solutions for fractional Kirchhoff problems with logarithmic nonlinearity, Nonlinear Anal., 198 (2020), 111899. http://dx.doi.org/10.1016/j.na.2020.111899 doi: 10.1016/j.na.2020.111899
|
| [24] |
M. Xiang, V. Rǎdulescu, B. Zhang, Existence results for singular fractional p-Kirchhoff problems, Acta Math. Sci., 42 (2022), 1209–1224. http://dx.doi.org/10.1007/s10473-022-0323-5 doi: 10.1007/s10473-022-0323-5
|
| 1. | Yuhai Zhao, Walter J. Lukiw, Microbiome-Mediated Upregulation of MicroRNA-146a in Sporadic Alzheimer’s Disease, 2018, 9, 1664-2295, 10.3389/fneur.2018.00145 | |
| 2. | Ting Pan, Han Shen, Songtao Yuan, Guohua Lu, Yi Zhang, Hanxue Wang, Yazhi Zhao, Xincheng Sun, Qinghuai Liu, Combined Transplantation With Human Mesenchymal Stem Cells Improves Retinal Rescue Effect of Human Fetal RPE Cells in Retinal Degeneration Mouse Model, 2020, 61, 1552-5783, 9, 10.1167/iovs.61.8.9 | |
| 3. | Leonid Minasyan, Parameswaran G. Sreekumar, David R. Hinton, Ram Kannan, Protective Mechanisms of the Mitochondrial-Derived Peptide Humanin in Oxidative and Endoplasmic Reticulum Stress in RPE Cells, 2017, 2017, 1942-0900, 1, 10.1155/2017/1675230 | |
| 4. | Ryoji Yanai, Shang Chen, Sho-Hei Uchi, Tomoaki Nanri, Kip M. Connor, Kazuhiro Kimura, Alfred S Lewin, Attenuation of choroidal neovascularization by dietary intake of ω-3 long-chain polyunsaturated fatty acids and lutein in mice, 2018, 13, 1932-6203, e0196037, 10.1371/journal.pone.0196037 | |
| 5. | Oksana Kutsyr, Xavier Sánchez-Sáez, Natalia Martínez-Gil, Emilio de Juan, Pedro Lax, Victoria Maneu, Nicolás Cuenca, Gradual Increase in Environmental Light Intensity Induces Oxidative Stress and Inflammation and Accelerates Retinal Neurodegeneration, 2020, 61, 1552-5783, 1, 10.1167/iovs.61.10.1 | |
| 6. | Andrea Maugeri, Martina Barchitta, Maria Mazzone, Francesco Giuliano, Guido Basile, Antonella Agodi, Resveratrol Modulates SIRT1 and DNMT Functions and Restores LINE-1 Methylation Levels in ARPE-19 Cells under Oxidative Stress and Inflammation, 2018, 19, 1422-0067, 2118, 10.3390/ijms19072118 | |
| 7. | Liria Yamamoto-Rodríguez, Marco A. Zarbin, Ricardo P. Casaroli-Marano, New frontiers and clinical implications in the pathophysiology of age-related macular degeneration, 2020, 154, 23870206, 496, 10.1016/j.medcle.2020.01.004 | |
| 8. | D. V. Telegina, O. S. Kozhevnikova, N. G. Kolosova, Changes in Retinal Glial Cells with Age and during Development of Age-Related Macular Degeneration, 2018, 83, 0006-2979, 1009, 10.1134/S000629791809002X | |
| 9. | Lindiwe M. Dlamini, Charlotte M. Tata, Marthe Carine F. Djuidje, Monisola I. Ikhile, Galina D. Nikolova, Yana D. Karamalakova, Veselina G. Gadjeva, Antoanetta M. Zheleva, Patrick B. Njobeh, Derek T. Ndinteh, Antioxidant and prooxidant effects of Piptadeniastrum africanum as the possible rationale behind its broad scale application in African ethnomedicine, 2019, 231, 03788741, 429, 10.1016/j.jep.2018.11.039 | |
| 10. | Yu Su, Yuexiong Yi, Lu Li, Changzheng Chen, circRNA-miRNA-mRNA network in age-related macular degeneration: From construction to identification, 2021, 203, 00144835, 108427, 10.1016/j.exer.2020.108427 | |
| 11. | Yeqi Zhou, Linbin Zhou, Kewen Zhou, Jingyue Zhang, Fu Shang, Xinyu Zhang, Celastrol Protects RPE Cells from Oxidative Stress-Induced Cell Death via Activation of Nrf2 Signaling Pathway, 2019, 19, 15665240, 172, 10.2174/1566524019666190424131704 | |
| 12. | Adrian Will-Orrego, Yubin Qiu, Elizabeth S. Fassbender, Siyuan Shen, Jorge Aranda, Namrata Kotagiri, Michael Maker, Sha-Mei Liao, Bruce D. Jaffee, Stephen H. Poor, Amount of Mononuclear Phagocyte Infiltrate Does Not Predict Area of Experimental Choroidal Neovascularization (CNV), 2018, 34, 1080-7683, 489, 10.1089/jop.2017.0131 | |
| 13. | Luigi Donato, Concetta Scimone, Giacomo Nicocia, Rosalia D’Angelo, Antonina Sidoti, RETRACTED ARTICLE: Role of oxidative stress in Retinitis pigmentosa: new involved pathways by an RNA-Seq analysis, 2019, 18, 1538-4101, 84, 10.1080/15384101.2018.1558873 | |
| 14. | Quiqing Zhao, Mingli Ji, Xuemei Wang, IL-10 inhibits retinal pigment epithelium cell proliferation and migration through regulation of VEGF in rhegmatogenous retinal detachment, 2018, 1791-2997, 10.3892/mmr.2018.8787 | |
| 15. | Kun Liu, Junwei Fang, Jing Jin, Shaopin Zhu, Xiaoyin Xu, Yupeng Xu, Bin Ye, Shu-Hai Lin, Xun Xu, Serum Metabolomics Reveals Personalized Metabolic Patterns for Macular Neovascular Disease Patient Stratification, 2020, 19, 1535-3893, 699, 10.1021/acs.jproteome.9b00574 | |
| 16. | Ruichan Li, Yanli Liu, Jing Xie, Xudong Huang, Li Zhang, Hua Liu, Lihua Li, Sirt3 mediates the protective effect of hydrogen in inhibiting ROS-induced retinal senescence, 2019, 135, 08915849, 116, 10.1016/j.freeradbiomed.2019.02.005 | |
| 17. | Maria Oltra, Lorena Vidal‐Gil, Rosa Maisto, Javier Sancho‐Pelluz, Jorge M. Barcia, Oxidative stress‐induced angiogenesis is mediated by miR‐205‐5p, 2020, 24, 1582-1838, 1428, 10.1111/jcmm.14822 | |
| 18. | Preeti Subramanian, S. Patricia Becerra, 2019, Chapter 62, 978-3-030-27377-4, 377, 10.1007/978-3-030-27378-1_62 | |
| 19. | Mala Upadhyay, Caroline Milliner, Brent A. Bell, Vera L. Bonilha, Oxidative stress in the retina and retinal pigment epithelium (RPE): Role of aging, and DJ-1, 2020, 37, 22132317, 101623, 10.1016/j.redox.2020.101623 | |
| 20. | Vibhuti Agrahari, Vivek Agrahari, Abhirup Mandal, Dhananjay Pal, Ashim K. Mitra, How are we improving the delivery to back of the eye? Advances and challenges of novel therapeutic approaches, 2017, 14, 1742-5247, 1145, 10.1080/17425247.2017.1272569 | |
| 21. | Maribel Vazquez, Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina, 2020, 11, 2072-666X, 1089, 10.3390/mi11121089 | |
| 22. | Suet Ding, Suresh Kumar, Pooi Mok, Cellular Reparative Mechanisms of Mesenchymal Stem Cells for Retinal Diseases, 2017, 18, 1422-0067, 1406, 10.3390/ijms18081406 | |
| 23. | Meshal Alshamrani, Sadia Sikder, Fohona Coulibaly, Abhirup Mandal, Dhananjay Pal, Ashim K. Mitra, Self-Assembling Topical Nanomicellar Formulation to Improve Curcumin Absorption Across Ocular Tissues, 2019, 20, 1530-9932, 10.1208/s12249-019-1404-1 | |
| 24. | 桥生 刘, Advance in Risk Factors and Prevention of Dry Age-Related Macular Degeneration, 2018, 07, 2167-6542, 21, 10.12677/HJO.2018.71004 | |
| 25. | The Redox Role of G6PD in Cell Growth, Cell Death, and Cancer, 2019, 8, 2073-4409, 1055, 10.3390/cells8091055 | |
| 26. | Sophie C. Lee, Steven Tran, Aana Amin, Lawrence S. Morse, Ala Moshiri, Susanna S. Park, Glenn Yiu, Retinal Vessel Density in Exudative and Nonexudative Age-Related Macular Degeneration on Optical Coherence Tomography Angiography, 2020, 212, 00029394, 7, 10.1016/j.ajo.2019.11.031 | |
| 27. | Ramazan Kürşad Zor, Serpil Erşan, Erkut Küçük, Gamze Yıldırım, İsmail Sarı, Serum malondialdehyde, monocyte chemoattractant protein-1, and vitamin C levels in wet type age-related macular degeneration patients, 2020, 12, 2515-8414, 251584142095168, 10.1177/2515841420951682 | |
| 28. | Pilar Herrero-Foncubierta, Jose Paredes, Maria Giron, Rafael Salto, Juan Cuerva, Delia Miguel, Angel Orte, A Red-Emitting, Multidimensional Sensor for the Simultaneous Cellular Imaging of Biothiols and Phosphate Ions, 2018, 18, 1424-8220, 161, 10.3390/s18010161 | |
| 29. | Andrea Maugeri, Martina Barchitta, Matteo Fallico, Niccolò Castellino, Michele Reibaldi, Antonella Agodi, Characterization of SIRT1/DNMTs Functions and LINE-1 Methylation in Patients with Age-Related Macular Degeneration, 2019, 8, 2077-0383, 159, 10.3390/jcm8020159 | |
| 30. | Andrea R. Waksmunski, Robert P. Igo, Yeunjoo E. Song, Jessica N. Cooke Bailey, Renee Laux, Denise Fuzzell, Sarada Fuzzell, Larry D. Adams, Laura Caywood, Michael Prough, Dwight Stambolian, William K. Scott, Margaret A. Pericak-Vance, Jonathan L. Haines, Rare variants and loci for age-related macular degeneration in the Ohio and Indiana Amish, 2019, 138, 0340-6717, 1171, 10.1007/s00439-019-02050-4 | |
| 31. | Maarten P. Rozing, Jon A. Durhuus, Marie Krogh Nielsen, Yousif Subhi, Thomas BL. Kirkwood, Rudi GJ. Westendorp, Torben Lykke Sørensen, Age-related macular degeneration: A two-level model hypothesis, 2020, 76, 13509462, 100825, 10.1016/j.preteyeres.2019.100825 | |
| 32. | Tarek K. Abouzed, Kadry M. Sadek, Mousa M. Ayoub, Ebeed A. Saleh, Sherif M. Nasr, Yasser S. El-Sayed, Moustafa Shoukry, Papaya extract upregulates the immune and antioxidants-related genes, and proteins expression in milk somatic cells of Friesian dairy cows, 2019, 103, 09312439, 407, 10.1111/jpn.13032 | |
| 33. | Alberto Modenese, Fabriziomaria Gobba, Macular degeneration and occupational risk factors: a systematic review, 2019, 92, 0340-0131, 1, 10.1007/s00420-018-1355-y | |
| 34. | Jasvinder A. Singh, John D. Cleveland, Der-Chong Tsai, Gout and the risk of age-related macular degeneration in the elderly, 2018, 13, 1932-6203, e0199562, 10.1371/journal.pone.0199562 | |
| 35. | Jae Yeon Kim, Sohae Park, So Hyun Park, Dongsook Lee, Gyu Hyun Kim, Jung Eun Noh, Kea Joo Lee, Gi Jin Kim, Overexpression of pigment epithelium-derived factor in placenta-derived mesenchymal stem cells promotes mitochondrial biogenesis in retinal cells, 2021, 101, 0023-6837, 51, 10.1038/s41374-020-0470-z | |
| 36. | Vibhuti Agrahari, Sulabh P. Patel, Nikhil Dhall, Zach Aulgur, Siddhant Thukral, Xiaoyan Yang, Ryan Conley, Ashim K. Mitra, Nanoparticles in thermosensitive gel based composite nanosystem for ocular diseases, 2018, 8, 2190-393X, 422, 10.1007/s13346-017-0435-y | |
| 37. | Jing Li, Jiaqi He, Xiang Zhang, Jiakai Li, Peiquan Zhao, Ping Fei, TSP1 ameliorates age-related macular degeneration by regulating the STAT3-iNOS signaling pathway, 2020, 388, 00144827, 111811, 10.1016/j.yexcr.2019.111811 | |
| 38. | Reem Hasaballah Alhasani, Lincoln Biswas, Ali Mohammad Tohari, Xinzhi Zhou, James Reilly, Jian-Feng He, Xinhua Shu, Gypenosides protect retinal pigment epithelium cells from oxidative stress, 2018, 112, 02786915, 76, 10.1016/j.fct.2017.12.037 | |
| 39. | Petra Schwarzer, Despina Kokona, Andreas Ebneter, Martin S. Zinkernagel, Effect of Inhibition of Colony-Stimulating Factor 1 Receptor on Choroidal Neovascularization in Mice, 2020, 190, 00029440, 412, 10.1016/j.ajpath.2019.10.011 | |
| 40. | Xiying Mao, Ting Pan, Han Shen, Huiyu Xi, Songtao Yuan, Qinghuai Liu, The rescue effect of mesenchymal stem cell on sodium iodate-induced retinal pigment epithelial cell death through deactivation of NF-κB-mediated NLRP3 inflammasome, 2018, 103, 07533322, 517, 10.1016/j.biopha.2018.04.038 | |
| 41. | Silvia Ravera, Alfonso Esposito, Paolo Degan, Federico Caicci, Lucia Manni, Anna Liguori, Angela Bisio, Valeria Iobbi, Anna Schito, Carlo Enrico Traverso, Isabella Panfoli, The diterpene Manool extracted from Salvia tingitana lowers free radical production in retinal rod outer segments by inhibiting the extramitochondrial F 1 F o ATP synthase , 2021, 0263-6484, 10.1002/cbf.3618 | |
| 42. | Arseny V Aybush, Alexander A Gulin, Alexander A Vasin, Alexander E Dontsov, Victor A Nadtochenko, Mikhail A Ostrovsky, Multimodal approach to reveal the effect of light irradiation on chemical composition of lipofuscin granules of human RPE tissues, 2020, 1695, 1742-6588, 012063, 10.1088/1742-6596/1695/1/012063 | |
| 43. | Sharareh Kalteh, Mostafa Saadat, Lack of association between three common genetic variations of XPC and susceptibility to age-related macular degeneration, a preliminary study, 2020, 21, 2090-2441, 10.1186/s43042-020-00060-w | |
| 44. | Maria L. Alonso-Alonso, Girish K. Srivastava, Ricardo Usategui-Martín, Maria T. García-Gutierrez, José Carlos Pastor, Ivan Fernandez-Bueno, Mesenchymal Stem Cell Secretome Enhancement by Nicotinamide and Vasoactive Intestinal Peptide: A New Therapeutic Approach for Retinal Degenerative Diseases, 2020, 2020, 1687-966X, 1, 10.1155/2020/9463548 | |
| 45. | Daniela Calzia, Paolo Degan, Federico Caicci, Maurizio Bruschi, Lucia Manni, Luca A. Ramenghi, Giovanni Candiano, Carlo Enrico Traverso, Isabella Panfoli, Modulation of the rod outer segment aerobic metabolism diminishes the production of radicals due to light absorption, 2018, 117, 08915849, 110, 10.1016/j.freeradbiomed.2018.01.029 | |
| 46. | Maiko Maruyama-Inoue, Shin Yamane, Hisayoshi Satoh, Shimpei Sato, Kazuaki Kadonosono, Choroidal Angioarchitecture According to Ultra-Widefield Indocyanine Green Angiography in Age-Related Macular Degeneration, 2017, 1, 2474-1264, 365, 10.1177/2474126417733161 | |
| 47. | Ting Zhang, Bobak Bahrami, Ling Zhu, 2018, Chapter 14, 978-3-319-89550-5, 273, 10.1007/978-3-319-89551-2_14 | |
| 48. | Virginia Puente-Muñoz, José M. Paredes, Sandra Resa, José Damaso Vílchez, Michal Zitnan, Delia Miguel, María Dolores Girón, Juan M. Cuerva, Rafael Salto, Luis Crovetto, New Thiol-Sensitive Dye Application for Measuring Oxidative Stress in Cell Cultures, 2019, 9, 2045-2322, 10.1038/s41598-018-38132-y | |
| 49. | Liria Yamamoto-Rodríguez, Marco A. Zarbin, Ricardo P. Casaroli-Marano, New frontiers and clinical implications in the pathophysiology of age-related macular degeneration, 2020, 154, 00257753, 496, 10.1016/j.medcli.2020.01.023 | |
| 50. | Vladimir Holan, Katerina Palacka, Barbora Hermankova, Mesenchymal Stem Cell-Based Therapy for Retinal Degenerative Diseases: Experimental Models and Clinical Trials, 2021, 10, 2073-4409, 588, 10.3390/cells10030588 | |
| 51. | Thomas J. Heesterbeek, Mansour Rouhi-Parkouhi, Stephanie J. Church, Yara T. Lechanteur, Laura Lorés-Motta, Nikolaos Kouvatsos, Simon J. Clark, Paul N. Bishop, Carel B. Hoyng, Anneke I. den Hollander, Richard D. Unwin, Anthony J. Day, Association of plasma trace element levels with neovascular age-related macular degeneration, 2020, 201, 00144835, 108324, 10.1016/j.exer.2020.108324 | |
| 52. | Reşat Duman, Ayhan Vurmaz, Role of innate immunity and oxidative stress in steroid-induced cataracts in developing chick embryos, 2018, 37, 1556-9527, 281, 10.1080/15569527.2018.1452929 | |
| 53. | R. M. Lucas, S. Yazar, A. R. Young, M. Norval, F. R. de Gruijl, Y. Takizawa, L. E. Rhodes, C. A. Sinclair, R. E. Neale, Human health in relation to exposure to solar ultraviolet radiation under changing stratospheric ozone and climate, 2019, 18, 1474-905X, 641, 10.1039/C8PP90060D | |
| 54. | Justin Hellman, Glenn Yiu, 2018, Chapter 2, 978-981-10-8544-4, 35, 10.1007/978-981-10-8545-1_2 | |
| 55. | Ting Zhang, Mark Gillies, Ying Wang, Weiyong Shen, Bobak Bahrami, Shaoxue Zeng, Meidong Zhu, Wenjuan Yao, Fanfan Zhou, Michael Murray, Ke Wang, Ling Zhu, Simvastatin protects photoreceptors from oxidative stress induced by all‐ trans ‐retinal, through the up‐regulation of interphotoreceptor retinoid binding protein , 2019, 176, 0007-1188, 2063, 10.1111/bph.14650 | |
| 56. | Yvette Wooff, Nilisha Fernando, Josephine H. C. Wong, Catherine Dietrich, Riemke Aggio-Bruce, Joshua A. Chu-Tan, Avril A. B. Robertson, Sarah L. Doyle, Si Ming Man, Riccardo Natoli, Caspase-1-dependent inflammasomes mediate photoreceptor cell death in photo-oxidative damage-induced retinal degeneration, 2020, 10, 2045-2322, 10.1038/s41598-020-58849-z | |
| 57. | Svetlana Trofimova, 2020, Chapter 1, 978-3-030-50159-4, 1, 10.1007/978-3-030-50160-0_1 | |
| 58. | Silvia Ravera, Alfonso Esposito, Paolo Degan, Federico Caicci, Daniela Calzia, Eleonora Perrotta, Lucia Manni, Angela Bisio, Valeria Iobbi, Anna Schito, Carlo Enrico Traverso, Isabella Panfoli, Sclareol modulates free radical production in the retinal rod outer segment by inhibiting the ectopic f1fo-atp synthase, 2020, 160, 08915849, 368, 10.1016/j.freeradbiomed.2020.08.014 | |
| 59. | Melissa K. Jones, Bin Lu, Dawn Zhaohui Chen, Weston R. Spivia, Augustus T. Mercado, Alexander V. Ljubimov, Clive N. Svendsen, Jennifer E. Eyk, Shaomei Wang, In Vitro and In Vivo Proteomic Comparison of Human Neural Progenitor Cell‐Induced Photoreceptor Survival, 2019, 19, 1615-9853, 1800213, 10.1002/pmic.201800213 | |
| 60. | Jiaowen Xu, Yuanyuan Tu, Ying Wang, Xun Xu, Xiaodong Sun, Laiqing Xie, Qingliang Zhao, Yang Guo, Yonghui Gu, Jingxia Du, Shu Du, Manhui Zhu, E. Song, Prodrug of epigallocatechin-3-gallate alleviates choroidal neovascularization via down-regulating HIF-1α/VEGF/VEGFR2 pathway and M1 type macrophage/microglia polarization, 2020, 121, 07533322, 109606, 10.1016/j.biopha.2019.109606 | |
| 61. | Luna Krstić, María J. González-García, Yolanda Diebold, Ocular Delivery of Polyphenols: Meeting the Unmet Needs, 2021, 26, 1420-3049, 370, 10.3390/molecules26020370 | |
| 62. | Myung Hee Kim, So Yeon Kwon, So-Yeun Woo, Woo Duck Seo, Dae Yu Kim, Antioxidative Effects of Chrysoeriol via Activation of the Nrf2 Signaling Pathway and Modulation of Mitochondrial Function, 2021, 26, 1420-3049, 313, 10.3390/molecules26020313 | |
| 63. | Silvia Ravera, Federico Caicci, Paolo Degan, Davide Maggi, Lucia Manni, Alessandra Puddu, Massimo Nicolò, Carlo E. Traverso, Isabella Panfoli, Inhibitory Action of Antidiabetic Drugs on the Free Radical Production by the Rod Outer Segment Ectopic Aerobic Metabolism, 2020, 9, 2076-3921, 1133, 10.3390/antiox9111133 | |
| 64. | Soo-Young Kim, Siva P. Kambhampati, Imran A. Bhutto, D. Scott McLeod, Gerard A. Lutty, Rangaramanujam M. Kannan, Evolution of oxidative stress, inflammation and neovascularization in the choroid and retina in a subretinal lipid induced age-related macular degeneration model, 2021, 203, 00144835, 108391, 10.1016/j.exer.2020.108391 | |
| 65. | Zofia Ulańczyk, Aleksandra Grabowicz, Elżbieta Cecerska-Heryć, Daria Śleboda-Taront, Elżbieta Krytkowska, Katarzyna Mozolewska-Piotrowska, Krzysztof Safranow, Miłosz Piotr Kawa, Barbara Dołęgowska, Anna Machalińska, Dietary and Lifestyle Factors Modulate the Activity of the Endogenous Antioxidant System in Patients with Age-Related Macular Degeneration: Correlations with Disease Severity, 2020, 9, 2076-3921, 954, 10.3390/antiox9100954 | |
| 66. | Vandana Soni, Vikas Pandey, Rahul Tiwari, Saket Asati, Rakesh K. Tekade, 2019, 9780128179093, 473, 10.1016/B978-0-12-817909-3.00013-3 | |
| 67. | Luis Fernando Hernández-Zimbrón, Ruben Zamora-Alvarado, Lenin Ochoa-De la Paz, Raul Velez-Montoya, Edgar Zenteno, Rosario Gulias-Cañizo, Hugo Quiroz-Mercado, Roberto Gonzalez-Salinas, Age-Related Macular Degeneration: New Paradigms for Treatment and Management of AMD, 2018, 2018, 1942-0900, 1, 10.1155/2018/8374647 | |
| 68. | Alberto Modenese, Leena Korpinen, Fabriziomaria Gobba, Solar Radiation Exposure and Outdoor Work: An Underestimated Occupational Risk, 2018, 15, 1660-4601, 2063, 10.3390/ijerph15102063 | |
| 69. | Saray Tabak, Sofia Schreiber-Avissar, Elie Beit-Yannai, Crosstalk between MicroRNA and Oxidative Stress in Primary Open-Angle Glaucoma, 2021, 22, 1422-0067, 2421, 10.3390/ijms22052421 | |
| 70. | Myung Hee Kim, Do-Hun Kim, Su Geun Yang, Dae Yu Kim, Improved effect of a mitochondria-targeted antioxidant on hydrogen peroxide-induced oxidative stress in human retinal pigment epithelium cells, 2021, 22, 2050-6511, 10.1186/s40360-020-00471-w | |
| 71. | José Carlos Rivera, Rabah Dabouz, Baraa Noueihed, Samy Omri, Houda Tahiri, Sylvain Chemtob, Ischemic Retinopathies: Oxidative Stress and Inflammation, 2017, 2017, 1942-0900, 1, 10.1155/2017/3940241 | |
| 72. | Sven Schnichels, François Paquet-Durand, Marina Löscher, Teresa Tsai, José Hurst, Stephanie C. Joachim, Alexa Klettner, Retina in a dish: Cell cultures, retinal explants and animal models for common diseases of the retina, 2021, 81, 13509462, 100880, 10.1016/j.preteyeres.2020.100880 | |
| 73. | Xiao Liu, Liwei Zhang, Jiang-Hui Wang, Huilan Zeng, Jingling Zou, Wei Tan, Han Zhao, Yan He, Jingming Shi, Shigeo Yoshida, Yunping Li, Yedi Zhou, Investigation of circRNA Expression Profiles and Analysis of circRNA-miRNA-mRNA Networks in an Animal (Mouse) Model of Age-Related Macular Degeneration, 2020, 45, 0271-3683, 1173, 10.1080/02713683.2020.1722179 | |
| 74. | Jie Bai, Yumei Yang, Dingting Wu, Fan Yang, SS‐31 protect retinal pigment epithelial cells from H 2 O 2 ‐induced cell injury by reducing apoptosis , 2021, 0305-1870, 10.1111/1440-1681.13484 | |
| 75. | Min Jae Ju, Junghoon Kim, Sung Kyun Park, Dong Hyun Kim, Yoon-Hyeong Choi, Long-term exposure to ambient air pollutants and age-related macular degeneration in middle-aged and older adults, 2022, 204, 00139351, 111953, 10.1016/j.envres.2021.111953 | |
| 76. | NESTAN MERKVILADZE, NIKOLOZ OBOLASHVILI, TAMAR MAISURADZE, MOLECULAR ASPECTS OF SOME RETINAL DISEASES, 2022, 15120392, 10.52340/jecm.2022.07.15 | |
| 77. | Lavinia Carlini, Gabriele Tancreda, Valeria Iobbi, Federico Caicci, Silvia Bruno, Alfonso Esposito, Daniela Calzia, Stefano Benini, Angela Bisio, Lucia Manni, Anna Schito, Carlo Enrico Traverso, Silvia Ravera, Isabella Panfoli, The Flavone Cirsiliol from Salvia x jamensis Binds the F1 Moiety of ATP Synthase, Modulating Free Radical Production, 2022, 11, 2073-4409, 3169, 10.3390/cells11193169 | |
| 78. | Charles C. Wykoff, Vrinda Hershberger, David Eichenbaum, Erin Henry, Husam S. Younis, Priya Chandra, Nancy Yuan, Mark Solloway, Alex DePaoli, Inhibition of Complement Factor 3 in Geographic Atrophy with NGM621: Phase 1 Dose-Escalation Study Results, 2022, 235, 00029394, 131, 10.1016/j.ajo.2021.08.018 | |
| 79. | Minghui Wang, Won-min Song, Chen Ming, Qian Wang, Xianxiao Zhou, Peng Xu, Azra Krek, Yonejung Yoon, Lap Ho, Miranda E. Orr, Guo-Cheng Yuan, Bin Zhang, Guidelines for bioinformatics of single-cell sequencing data analysis in Alzheimer’s disease: review, recommendation, implementation and application, 2022, 17, 1750-1326, 10.1186/s13024-022-00517-z | |
| 80. | Victoria Maneu, Pedro Lax, Antonio Miguel G. De Diego, Nicolás Cuenca, Antonio G. García, Combined drug triads for synergic neuroprotection in retinal degeneration, 2022, 149, 07533322, 112911, 10.1016/j.biopha.2022.112911 | |
| 81. | Alberto Melecchi, Rosario Amato, Dominga Lapi, Massimo Dal Monte, Dario Rusciano, Paola Bagnoli, Maurizio Cammalleri, Increased efficacy of dietary supplement containing wax ester-rich marine oil and xanthophylls in a mouse model of dry macular degeneration, 2022, 13, 1663-9812, 10.3389/fphar.2022.1038730 | |
| 82. | Alexander Wade, Rameshu Rallabandi, Steven Lucas, Catrina Oberg, Aruna Gorusupudi, Paul S. Bernstein, Jon D. Rainier, The synthesis of the very long chain polyunsaturated fatty acid (VLC-PUFA) 32:6 n-3, 2021, 19, 1477-0520, 5563, 10.1039/D1OB00491C | |
| 83. | Fátima Milhano dos Santos, Sergio Ciordia, Joana Mesquita, João Paulo Castro de Sousa, Alberto Paradela, Cândida Teixeira Tomaz, Luís António Paulino Passarinha, Vitreous humor proteome: unraveling the molecular mechanisms underlying proliferative and neovascular vitreoretinal diseases, 2023, 80, 1420-682X, 10.1007/s00018-022-04670-y | |
| 84. | R. M. Lucas, S. Yazar, A. R. Young, M. Norval, F. R. de Gruijl, Y. Takizawa, L. E. Rhodes, C. A. Sinclair, R. E. Neale, Human health in relation to exposure to solar ultraviolet radiation under changing stratospheric ozone and climate, 2019, 18, 1474-905X, 641, 10.1039/c8pp90060d | |
| 85. | Justin C. Muste, Matthew W. Russell, Andrew X. Chen, Kanika Seth, Amogh I. Iyer, Carolina C. S. Valentim, Anna K. Wu, Blanche L. Kuo, Aneesha Kalur, Resya Sastry, Grant L. Hom, Thais F. Conti, Collin A. Rich, Katherine E. Talcott, Sumit Sharma, Rishi P. Singh, Functional Imaging of Mitochondria in Age-Related Macular Degeneration Using Flavoprotein Fluorescence, 2023, 54, 2325-8160, 24, 10.3928/23258160-20221214-03 | |
| 86. | Jun Wang, Mengling Li, Ziyue Geng, Saadullah Khattak, Xinying Ji, Dongdong Wu, Yalong Dang, Francisco Rios, Role of Oxidative Stress in Retinal Disease and the Early Intervention Strategies: A Review, 2022, 2022, 1942-0994, 1, 10.1155/2022/7836828 | |
| 87. | Zhongjie Fu, Raffael Liegl, Zhongxiao Wang, Yan Gong, Chi-Hsiu Liu, Ye Sun, Bertan Cakir, Samuel B. Burnim, Steven S. Meng, Chatarina Löfqvist, John Paul SanGiovanni, Ann Hellström, Lois E. H. Smith, Adiponectin Mediates Dietary Omega-3 Long-Chain Polyunsaturated Fatty Acid Protection Against Choroidal Neovascularization in Mice, 2017, 58, 1552-5783, 3862, 10.1167/iovs.17-21796 | |
| 88. | Kristin Krueger, Elke Boehme, Alexa Karina Klettner, Marietta Zille, The potential of marine resources for retinal diseases: a systematic review of the molecular mechanisms, 2022, 62, 1040-8398, 7518, 10.1080/10408398.2021.1915242 | |
| 89. | Ayat Mahmoud Domouky, Walaa M. Samy, Walaa A. Rashad, Therapeutic effect of the mesenchymal stem cells on vigabatrin-induced retinopathy in adult male albino rat, 2022, 55, 2093-3665, 217, 10.5115/acb.22.006 | |
| 90. | Zhijie Wang, Yinhua Huang, Feixue Chu, Kai Liao, Zekai Cui, Jiansu Chen, Shibo Tang, Integrated Analysis of DNA methylation and transcriptome profile to identify key features of age-related macular degeneration, 2021, 12, 2165-5979, 7061, 10.1080/21655979.2021.1976502 | |
| 91. | Marina Yakovleva, Alexander Dontsov, Natalia Trofimova, Natalia Sakina, Alexey Kononikhin, Arseny Aybush, Alexander Gulin, Tatiana Feldman, Mikhail Ostrovsky, Lipofuscin Granule Bisretinoid Oxidation in the Human Retinal Pigment Epithelium forms Cytotoxic Carbonyls, 2021, 23, 1422-0067, 222, 10.3390/ijms23010222 | |
| 92. | Kathleen Romond, Minhaj Alam, Sasha Kravets, Luis de Sisternes, Theodore Leng, Jennifer I Lim, Daniel Rubin, Joelle A Hallak, Imaging and artificial intelligence for progression of age-related macular degeneration, 2021, 246, 1535-3702, 2159, 10.1177/15353702211031547 | |
| 93. | Fukutaro Mano, Shoei Sakata, Kuo-Chung Chang, Tomiya Mano, Effects of Zinc Acetate Hydrate Treatment on Serum Oxidative Stress Markers in Patients with Macular Drusen, 2021, 37, 1080-7683, 518, 10.1089/jop.2021.0031 | |
| 94. | Shan-Shan Li, Hui-Hui Wang, Dawei Zhang, Efficacy of different nutrients in age-related macular degeneration: A systematic review and network meta-analysis, 2022, 37, 0882-0538, 515, 10.1080/08820538.2021.2022165 | |
| 95. | Alexa Klettner, Johann Roider, Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration, 2021, 22, 1422-0067, 8387, 10.3390/ijms22168387 | |
| 96. | Julia Hildebrandt, Tom Käckenmeister, Katrin Winkelmann, Philipp Dörschmann, Johann Roider, Alexa Klettner, Pro-inflammatory activation changes intracellular transport of bevacizumab in the retinal pigment epithelium in vitro, 2022, 260, 0721-832X, 857, 10.1007/s00417-021-05443-2 | |
| 97. | Malgorzata Mrowicka, Jerzy Mrowicki, Ewa Kucharska, Barbara Smigielska, Jacek Pawel Szaflik, Jerzy Szaflik, Ireneusz Majsterek, The Role of Oxidative Stress and the Importance of miRNAs as Potential Biomarkers in the Development of Age-Related Macular Degeneration, 2021, 9, 2227-9717, 1328, 10.3390/pr9081328 | |
| 98. | Iswariyaraja Sridevi Gurubaran, Hanna Heloterä, Stephen Marry, Ali Koskela, Juha M. T. Hyttinen, Jussi J. Paterno, Arto Urtti, Mei Chen, Heping Xu, Anu Kauppinen, Kai Kaarniranta, Oxidative Stress and Mitochondrial Damage in Dry Age-Related Macular Degeneration Like NFE2L2/PGC-1α -/- Mouse Model Evoke Complement Component C5a Independent of C3, 2021, 10, 2079-7737, 622, 10.3390/biology10070622 | |
| 99. | Kah-Hui Wong, Hui-Yin Nam, Sze-Yuen Lew, Murali Naidu, Pamela David, Tengku Ain Kamalden, Siti Nurma Hanim Hadie, Lee-Wei Lim, Discovering the Potential of Natural Antioxidants in Age-Related Macular Degeneration: A Review, 2022, 15, 1424-8247, 101, 10.3390/ph15010101 | |
| 100. | Heran Getachew, Blanca Chinchilla, Rosario Fernandez-Godino, 2021, Chapter 409, 978-1-0716-2584-2, 321, 10.1007/7651_2021_409 | |
| 101. | Philipp Dörschmann, Hubeydullah Akkurt, Georg Kopplin, Maria Dalgaard Mikkelsen, Anne S. Meyer, Johann Roider, Alexa Klettner, Establishment of specific age-related macular degeneration relevant gene expression panels using porcine retinal pigment epithelium for assessing fucoidan bioactivity, 2023, 231, 00144835, 109469, 10.1016/j.exer.2023.109469 | |
| 102. | Philipp Dörschmann, Charlotte Seeba, Tabea Thalenhorst, Johann Roider, Alexa Klettner, Anti-inflammatory properties of antiangiogenic fucoidan in retinal pigment epithelium cells, 2023, 9, 24058440, e15202, 10.1016/j.heliyon.2023.e15202 | |
| 103. | Samantha Sasseville, Samira Karami, Ange Tchatchouang, Pascale Charpentier, Princia Anney, Delphine Gobert, Stéphanie Proulx, Biomaterials used for tissue engineering of barrier-forming cell monolayers in the eye, 2023, 11, 2296-4185, 10.3389/fbioe.2023.1269385 | |
| 104. | Bryanna Lee, Natalie A. Afshari, Peter X. Shaw, Oxidative stress and antioxidants in cataract development, 2024, 35, 1040-8738, 57, 10.1097/ICU.0000000000001009 | |
| 105. | Sijie Gu, Siqi Wu, Zesong Lin, Zhuo Han, Kunlun Mo, Huaxing Huang, Mingsen Li, Gen Li, Hong Ouyang, Li Wang, Screening and evaluation of antioxidants for retinal pigment epithelial cell protection: L-ergothioneine as a novel therapeutic candidate through NRF2 activation, 2024, 242, 00144835, 109862, 10.1016/j.exer.2024.109862 | |
| 106. | Silvia Ravera, Nadia Bertola, Alessandra Puddu, Silvia Bruno, Davide Maggi, Isabella Panfoli, Crosstalk between the Rod Outer Segments and Retinal Pigmented Epithelium in the Generation of Oxidative Stress in an In Vitro Model, 2023, 12, 2073-4409, 2173, 10.3390/cells12172173 | |
| 107. | 2023, 21, 13040855, 10.6002/ect.2021.0360 | |
| 108. | Dong Hyun Jo, Su Hyun Lee, Minsol Jeon, Chang Sik Cho, Da-Eun Kim, Hyunkyung Kim, Jeong Hun Kim, Activation of Lysosomal Function Ameliorates Amyloid-β-Induced Tight Junction Disruption in the Retinal Pigment Epithelium, 2023, 46, 10168478, 675, 10.14348/molcells.2023.0056 | |
| 109. | Suresh Khadke, Poonam Gupte, Akanksha Mourya, Amit Yadav, Sarika Mane, Asavari Joshi, Madhavi Mahajan, Manisha Mishra, Supriya Bhalerao, Immunomodulatory effect of a proprietary polyherbal formulation on healthy participants: A single- blind, randomized, placebo- controlled, exploratory clinical study, 2023, 14, 2229-3485, 130, 10.4103/picr.picr_100_22 | |
| 110. | Sandeep M. Subrahmanian, Esma I. Yerlikaya, Siddharth Sunilkumar, Allyson L. Toro, Christopher M. McCurry, Stephanie L. Grillo, Alistair J. Barber, Jeffrey M. Sundstrom, Michael D. Dennis, Deletion of the stress response protein REDD1 prevents sodium iodate-induced RPE damage and photoreceptor loss, 2024, 2509-2723, 10.1007/s11357-024-01362-2 | |
| 111. | Małgorzata B. Różanowska, Lipofuscin, Its Origin, Properties, and Contribution to Retinal Fluorescence as a Potential Biomarker of Oxidative Damage to the Retina, 2023, 12, 2076-3921, 2111, 10.3390/antiox12122111 | |
| 112. | Philipp Dörschmann, Tabea Thalenhorst, Charlotte Seeba, Marie-Theres Tischhöfer, Sandesh Neupane, Johann Roider, Susanne Alban, Alexa Klettner, Comparison of Fucoidans from Saccharina latissima Regarding Age-Related Macular Degeneration Relevant Pathomechanisms in Retinal Pigment Epithelium, 2023, 24, 1422-0067, 7939, 10.3390/ijms24097939 | |
| 113. | Alfar Ahamed, Rendy Hosea, Shourong Wu, Vivi Kasim, The Emerging Roles of the Metabolic Regulator G6PD in Human Cancers, 2023, 24, 1422-0067, 17238, 10.3390/ijms242417238 | |
| 114. | Yao Wang, Xianning Liu, Bei Wang, Hanhan Sun, Yiqian Ren, Hongbing Zhang, Compounding engineered mesenchymal stem cell-derived exosomes: A potential rescue strategy for retinal degeneration, 2024, 173, 07533322, 116424, 10.1016/j.biopha.2024.116424 | |
| 115. | Menno E. Sluijter, Alexandre Teixeira, Kris Vissers, Luis Josino Brasil, Bert van Duijn, The Anti-Inflammatory Action of Pulsed Radiofrequency—A Hypothesis and Potential Applications, 2023, 11, 2076-3271, 58, 10.3390/medsci11030058 | |
| 116. | Jae Yong Park, Jae Suk Kim, Ha Eun Sim, Seung Hyun Lee, Hyun Min Na, Min Ji Kang, Je Hyung Hwang, Prevalence and Risk Factors of Age-Related Macular Degeneration Features Among Pilots of the Republic of Korea Air Force, 2023, 0275-004X, 10.1097/IAE.0000000000003976 | |
| 117. | Iswariyaraja Sridevi Gurubaran, Mitochondrial damage and clearance in retinal pigment epithelial cells, 2024, 102, 1755-375X, 3, 10.1111/aos.16661 | |
| 118. | Dao Nguyen, Thilini Thrimawithana, Terrence J. Piva, Danilla Grando, Tien Huynh, Benefits of plant carotenoids against age-related macular degeneration, 2023, 106, 17564646, 105597, 10.1016/j.jff.2023.105597 | |
| 119. | Anindya Samanta, Amer F. Alsoudi, Ehsan Rahimy, Jay Chhablani, Christina Y. Weng, Imaging Modalities for Dry Macular Degeneration, 2024, 64, 0020-8167, 35, 10.1097/IIO.0000000000000512 | |
| 120. | Maya Carleton, Nicholas W. Oesch, Bridging the gap of vision restoration, 2024, 18, 1662-5102, 10.3389/fncel.2024.1502473 | |
| 121. | Alessandra Puddu, Massimo Nicolò, Davide C. Maggi, Combination of Saffron (Crocus sativus), Elderberry (Sambucus nigra L.) and Melilotus officinalis Protects ARPE-19 Cells from Oxidative Stress, 2025, 26, 1422-0067, 1496, 10.3390/ijms26041496 | |
| 122. | Kun‐Lin Yeh, Yu‐Hsiang Kuan, Sheng‐Wen Wu, Chen‐Yu Chiang, Chun‐Jung Chen, Wen‐Ying Chen, Chi‐Chung Chou, Irigenin Alleviates Blue Light‐Induced Retinal Damage by Upregulating Antioxidative Defense System via Nrf2 Pathway In Vivo and In Vitro, 2025, 1520-4081, 10.1002/tox.24501 | |
| 123. | Chávez García Ricardo, Ortega Camarillo Clara, Contreras Ramos Alejandra, Díaz Rosas Guadalupe, Avalos Rodríguez Alejandro, C.C.J.R. Pérez-Rivero, Vergara Onofre Marcela, Comparison of G6PD activity with a modified hormonal environment during the critical period of hypothalamic sexual differentiation in Sprague Dawley rats, 2025, 03043940, 138206, 10.1016/j.neulet.2025.138206 |

Zusheng Chen, Hongmin Suo, Jun Lei. Multiple solutions for a fractional p-Kirchhoff equation with critical growth and low order perturbations[J]. AIMS Mathematics, 2022, 7(7): 12897-12912. doi: 10.3934/math.2022714

| Genes | Variants (SNP) | Full Name | Function | Position | Odds Ratios | References |
| ABCA4 | rs1800553, rs1800555 | ATP-binding cassette, sub-family A (ABC1), member 4 | Photoreceptor specific expression; transport of N-retinylidene-PE across the outer segment disc membrane | 1p22 | N.A. | [10,11,12] |
| APOE | rs429358, rs7412 | apolipoprotein E | lipid and cholesterol transport | 19q13 | ε2 ORhomo = 1.046; ε4 ORhomo = 0.847 (Thakkinstian); ε4 ORhomo = 0.35-0.53, 0.847 (separate report-Francis) | [13,14] |
| ARMS2 /LOC387715* | rs10490924, in/del (372_815delins54) | age-related maculopathy susceptibility 2 | no known function; protein localized to mitochondrial outer membrane | 10q26 | ORhomo = 8.59 (rs10490924) | [12,15,16,17] |
| HTRA1* | rs11200638 | human High Temperature Requirement A1 | trypsin-like serine protease | 10q26 | ORhomo = 6.92, 7.46 | [6,18,19] |
| C2/CFB | rs9332739 (c2), rs4151667(CFB), rs641153 (CFB) | complement 2/ complement factor B | regulation of complement activation | 6p21 | ORhetero = 0.21-0.45 | [6,20] |
| C3 | rs2230199, rs1047286 | complement 3 | Innate immunity (alternative complement pathway activator, classical pathway component) | 19p13 | ORhomo = 1.93-3.26 | [21,22,23] |
| CETP | rs1864163 | Cholesteryl ester transfer protein | Transfer cholesteryl esters between lipoproteins | 16q21 | ORhomo = 1.2 | [24] |
| CFH* | rs1061170 | complement factor H | inhibitor of alternative complement pathway | 1q32 | ORhomo = 6.35 | [25,26,27,28] |
| CFHR1 /CFHR3 | 84K bp deletion | complement factor H-related 1, 3 | exact function unknown, possible overlapping function with CFH | 1q31-q32 | ORhomo = 0.29 | [29,30] |
| CFI | rs4698775 | Complement factor I | regulation of complement activation | 4q25 | ORhomo = 1.1 | [31] |
| CX3CR1 | rs3732378 | chemokine (C-X3-C motif) receptor 1 | Inflammatory (chemokine receptor) | 3p21 | ORhomo = 1.98-2.70 | [32,33] |
| ERCC6 | rs3793784 | excision-repain cross-complementing, group6 | DNA repair | 10q11 | ORhomo = 1.6 | [34] |
| LIPC | rs920915 | Hepatic lipase | Lipid metabolism | 15q21-23 | ORhetero = 1.1 | [24,35/a>,36] |
| TIMP3 | rs9621532 | Tissue inhibitor of metalloproteinase | Complexes with inactive metalloproteinase | 22q12 | ORhomo = 1.31-1.91 | [37,38,39] |
| TLR3 | rs3775291 | toll-like receptor 3 | Innate immunity (encodes protein to recognize viral dsRNA) | 4q35 | ORhomo = 0.44-0.61 | [40,41] |
| TLR4 | rs4986790 | toll-like receptor 4 | Innate immunity (encodes bacterial endotoxin receptor) | 9q32-q33 | ORhetero = 2.65 | [32,42,43] |
| VEGFA | rs833069, rs1413711 | vascular endothelial growth factor A | Angiogenic and vasculogenic growth factor | 6p12 | ORhomo = 5.29 (rs833069), ORhomo = 2.40 (rs1413711) | [44,45,46] |
| *ORhomo indicates odds ratio from homozygous; ORhetero indicates odds ratio of heterozygous. | ||||||
DownLoad: CSV| Genes | Variants (SNP) | Full Name | Function | Position | Odds Ratios | References |
| ABCA4 | rs1800553, rs1800555 | ATP-binding cassette, sub-family A (ABC1), member 4 | Photoreceptor specific expression; transport of N-retinylidene-PE across the outer segment disc membrane | 1p22 | N.A. | [10,11,12] |
| APOE | rs429358, rs7412 | apolipoprotein E | lipid and cholesterol transport | 19q13 | ε2 ORhomo = 1.046; ε4 ORhomo = 0.847 (Thakkinstian); ε4 ORhomo = 0.35-0.53, 0.847 (separate report-Francis) | [13,14] |
| ARMS2 /LOC387715* | rs10490924, in/del (372_815delins54) | age-related maculopathy susceptibility 2 | no known function; protein localized to mitochondrial outer membrane | 10q26 | ORhomo = 8.59 (rs10490924) | [12,15,16,17] |
| HTRA1* | rs11200638 | human High Temperature Requirement A1 | trypsin-like serine protease | 10q26 | ORhomo = 6.92, 7.46 | [6,18,19] |
| C2/CFB | rs9332739 (c2), rs4151667(CFB), rs641153 (CFB) | complement 2/ complement factor B | regulation of complement activation | 6p21 | ORhetero = 0.21-0.45 | [6,20] |
| C3 | rs2230199, rs1047286 | complement 3 | Innate immunity (alternative complement pathway activator, classical pathway component) | 19p13 | ORhomo = 1.93-3.26 | [21,22,23] |
| CETP | rs1864163 | Cholesteryl ester transfer protein | Transfer cholesteryl esters between lipoproteins | 16q21 | ORhomo = 1.2 | [24] |
| CFH* | rs1061170 | complement factor H | inhibitor of alternative complement pathway | 1q32 | ORhomo = 6.35 | [25,26,27,28] |
| CFHR1 /CFHR3 | 84K bp deletion | complement factor H-related 1, 3 | exact function unknown, possible overlapping function with CFH | 1q31-q32 | ORhomo = 0.29 | [29,30] |
| CFI | rs4698775 | Complement factor I | regulation of complement activation | 4q25 | ORhomo = 1.1 | [31] |
| CX3CR1 | rs3732378 | chemokine (C-X3-C motif) receptor 1 | Inflammatory (chemokine receptor) | 3p21 | ORhomo = 1.98-2.70 | [32,33] |
| ERCC6 | rs3793784 | excision-repain cross-complementing, group6 | DNA repair | 10q11 | ORhomo = 1.6 | [34] |
| LIPC | rs920915 | Hepatic lipase | Lipid metabolism | 15q21-23 | ORhetero = 1.1 | [24,35/a>,36] |
| TIMP3 | rs9621532 | Tissue inhibitor of metalloproteinase | Complexes with inactive metalloproteinase | 22q12 | ORhomo = 1.31-1.91 | [37,38,39] |
| TLR3 | rs3775291 | toll-like receptor 3 | Innate immunity (encodes protein to recognize viral dsRNA) | 4q35 | ORhomo = 0.44-0.61 | [40,41] |
| TLR4 | rs4986790 | toll-like receptor 4 | Innate immunity (encodes bacterial endotoxin receptor) | 9q32-q33 | ORhetero = 2.65 | [32,42,43] |
| VEGFA | rs833069, rs1413711 | vascular endothelial growth factor A | Angiogenic and vasculogenic growth factor | 6p12 | ORhomo = 5.29 (rs833069), ORhomo = 2.40 (rs1413711) | [44,45,46] |
| *ORhomo indicates odds ratio from homozygous; ORhetero indicates odds ratio of heterozygous. | ||||||





