Citation: Volodymyr Ivanov, Viktor Stabnikov, Olena Stabnikova, Anatoliy Salyuk, Evhenii Shapovalov, Zubair Ahmed, Joo Hwa Tay. Iron-containing clay and hematite iron ore in slurry-phase anaerobic digestion of chicken manure[J]. AIMS Materials Science, 2019, 6(5): 821-832. doi: 10.3934/matersci.2019.5.821

| [1] |
Andre L, Pauss A, Ribeiro T (2018) Solid anaerobic digestion: state-of-art, scientific and technological hurdles. Bioresource Technol 247: 1027–1037. doi: 10.1016/j.biortech.2017.09.003

|
| [2] |
Bujoczek G, Oleszkiewicz J, Sparling R, et al. (2000) High solid anaerobic digestion of chicken manure. J Agr Eng Res 76: 51–60. doi: 10.1006/jaer.2000.0529
|
| [3] |
Ge X, Xu F, Li Y (2016) Solid-state anaerobic digestion of lignocellulosic biomass: recent progress and perspectives. Bioresource Technol 205: 239–249. doi: 10.1016/j.biortech.2016.01.050
|
| [4] |
Yang L, Xu F, Ge X, et al. (2015) Challenges and strategies for solid-state anaerobic digestion of lignocellulosic biomass. Renew Sust Energ Rev 44: 824–834. doi: 10.1016/j.rser.2015.01.002
|
| [5] |
Wei P, Mudde RF, Uijttewaal WSJ, et al. (2019) Characterising the two-phase flow and mixing performance in a gas-mixed anaerobic digester: importance for scaled-up applications. Water Res 149: 86–97. doi: 10.1016/j.watres.2018.10.077
|
| [6] |
Wang H, Tao Y, Temudo M, et al. (2015) An integrated approach for efficient biomethane production from solid bio-wastes in a compact system. Biotechnol Biofuels 8: 62. doi: 10.1186/s13068-015-0237-8
|
| [7] | Marks PJ, Wujcik WJ, Loncar AF (1994) Remediation Technologies Screening Matrix and Reference Guide, Version 4.0. Available from: https://frtr.gov/matrix2/section4/4-14.html. |
| [8] |
Vamini B, Vianney T, Jo YS (2017) Water for small-scale biogas digesters in Sub-Saharan Africa. GCB Bioenergy 9: 339–357. doi: 10.1111/gcbb.12339
|
| [9] |
Rajagopal R, Massé DI, Singh G (2013) A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresource Technol 143: 632–641. doi: 10.1016/j.biortech.2013.06.030
|
| [10] |
Yenigun O, Demirel B (2013) Ammonia inhibition in anaerobic digestion: a review. Process Biochem 48: 901–911. doi: 10.1016/j.procbio.2013.04.012
|
| [11] |
Niu Q, Qiao W, Qiang H, et al. (2013) Mesophilic methane fermentation of chicken manure at a wide range of ammonia concentration: stability, inhibition and recovery. Bioresource Technol 137: 358–367. doi: 10.1016/j.biortech.2013.03.080
|
| [12] |
Niu Q, Kubota K, Qiao W, et al. (2015) Effect of ammonia inhibition on microbial community dynamic and process functional resilience in mesophilic methane fermentation of chicken manure. J Chem Technol Biot 90: 2161–2169. doi: 10.1002/jctb.4527
|
| [13] | Salyuk AI, Zhadan SO, Shapovalov EB (2014) Thermophilic methane digestion of chicken manure. Ukrainian Food J 3: 587–594. |
| [14] | Salyuk AI, Zhadan SO, Shapovalov EB (2015) Thermophilic methane fermentation of chicken manure in a wide range of substrate moisture contents. J Food Packag Sci Tech Technol 4: 36–40. |
| [15] |
Zhang W, Lau A (2007) Reducing ammonia emission from poultry manure composting via struvite formation. J Chem Technol Biot 82: 598–602. doi: 10.1002/jctb.1701
|
| [16] |
Krakat N, Demirel B, Anjum R (2017) Methods of ammonia removal in anaerobic digestion: a review. Water Sci Technol 76: 1925–1938. doi: 10.2166/wst.2017.406
|
| [17] | Zhang L, Lee Y, Jahng D (2012) Ammonia stripping for enhanced biomethanization of piggery wastewater. J Hazard Mater 199: 36–42. |
| [18] |
Surmeli RO, Bayrakdar A, Calli B (2017) Removal and recovery of ammonia from chicken manure. Water Sci Technol 75: 2811–2817. doi: 10.2166/wst.2017.116
|
| [19] |
Markou G (2015) Improved anaerobic digestion performance and biogas production from poultry litter after lowering its nitrogen content. Bioresource Technol 196: 726–730. doi: 10.1016/j.biortech.2015.07.067
|
| [20] |
Abouelenien F, Fujiwara W, Namba Y, et al. (2010) Improved methane fermentation of chicken manure via ammonia removal by biogas recycle. Bioresource Technol 101: 6368–6373. doi: 10.1016/j.biortech.2010.03.071
|
| [21] |
Laureni M, Palatsi J, Llovera M, et al. (2013) Influence of pig slurry characteristics on ammonia stripping efficiencies and quality of the recovered ammonium-sulfate solution. J Chem Technol Biot 88: 1654–1662. doi: 10.1002/jctb.4016
|
| [22] |
Alshameri A, He H, Zhu J, et al. (2018) Adsorption of ammonium by different natural clay minerals: characterization, kinetics and adsorption isotherms. Appl Clay Sci 159: 83–93. doi: 10.1016/j.clay.2017.11.007
|
| [23] |
Zhu R, Chen Q, Zhou Q, et al. (2016) Adsorbents based on montmorillonite for contaminant removal from water: A review. Appl Clay Sci 123: 239–258. doi: 10.1016/j.clay.2015.12.024
|
| [24] |
Borisover M, Davis JA (2015) Adsorption of inorganic and organic solutes by clay minerals, In: Tournassat C, Steefel C, Bourg I, et al., Natural and Engineered Clay Barriers, Elsevier 6: 33–70. doi: 10.1016/B978-0-08-100027-4.00002-4
|
| [25] |
Khosravi A, Esmhosseini M, Khezri S (2014) Removal of ammonium ion from aqueous solutions using natural zeolite: kinetic, equilibrium and thermodynamic studies. Res Chem Intermediat 40: 2905–2917. doi: 10.1007/s11164-013-1137-9
|
| [26] |
Rožić M, Cerjan-Stefanovic S, Kurajica S, et al. (2000) Ammonical nitrogen removal from water by treatment with clays and zeolites. Water Res 34: 3675–3681. doi: 10.1016/S0043-1354(00)00113-5
|
| [27] | Ma JY, Pan JT, Gao TL, et al. (2016) Enhanced anaerobic digestion of chicken manure by bentonite addition. Res Environ Sci 29: 442–448. |
| [28] |
Chen H, Awasthi MK, Liu T, et al. (2018) Influence of clay as additive on greenhouse gases emission and maturity evaluation during chicken manure composting. Bioresource Technol 266: 82–88. doi: 10.1016/j.biortech.2018.06.073
|
| [29] |
Ivanov V, Stabnikov V, Guo CH, et al. (2014) Wastewater engineering applications of BioIronTech process based on the biogeochemical cycle of iron bioreduction and (bio)oxidation. AIMS Environ J 1: 53–66. doi: 10.3934/environsci.2014.2.53
|
| [30] |
Ivanov V, Stabnikov V, Tay JH (2018) Removal of the recalcitrant artificial sweetener sucralose and its by-products from industrial wastewater using microbial reduction/oxidation of iron. ChemEngineering 2: 37. doi: 10.3390/chemengineering2030037
|
| [31] |
Stabnikov VP, Tay STL, Tay JH, et al. (2004) Effect of iron hydroxide on phosphate removal during anaerobic digestion of activated sludge. Appl Biochem Micro 40: 376–380. doi: 10.1023/B:ABIM.0000033914.52026.e5
|
| [32] |
Binner I, Dultz S, Schellhorn M, et al. (2017) Potassium adsorption and release properties of clays in peat-based horticultural substrates for increasing the cultivation safety of plants. Appl Clay Sci 145: 28–36. doi: 10.1016/j.clay.2017.05.013
|
| [33] | Visser A, Nozhevnikova AN, Lettinga G (1993) Sulphide inhibition of methanogenic activity at various pH levels at 55 ℃. J Chem Technol Biot 57: 9–14. |
| [34] |
Koster IW, Rinzema A, De Vegt AL, et al. (1986) Sulfide inhibition of the methanogenic activity of granular sludge at various pH levels. Water Res 20: 1561–1567. doi: 10.1016/0043-1354(86)90121-1
|
| [35] | Muhlbauer RV, Swestka RJ, Burns RT, et al. (2008) Development and testing of a hydrogen sulfide detection system for use in swine housing. ASABE 6: 084203. |
| [36] | Occupational Safety and Health Administration (2005) Available from: https://www.osha.gov/SLTC/hydrogensulfide/hazards.html. |
| [37] | Yuzir A, Yaacob SS, Tijani H, et al. (2017) Addition of ferric chloride in anaerobic digesters to enhance sulphide removal and methanogenesis. Desalin Water Treat 79: 64–72. |
| [38] | Stabnikov VP, Ivanov VN (2006) The effect of various iron hydroxide concentrations on the anaerobic fermentation of sulfate-containing model wastewater. Appl Biochem Micro 42: 284–288. |
| [39] |
Stabnikov V, Ivanov V (2017) Biotechnological production of biogrout from iron ore and cellulose. J Chem Technol Biot 92: 180–187. doi: 10.1002/jctb.4989
|
| [40] | Stucki JW (2006) Properties and behaviour of iron in clay minerals, In: Bergaya F, Theng BKG, Lagaly G, Developments in Clay Science, Elsevier Science Ltd 1: 423–475. |
| [41] | Markos N (2003) Bentonite-iron interactions in natural occurrences and in laboratory-the effects of the interactions on the properties of bentonite: a literature survey. Working report 2003-55, Posiva Oy. |
| [42] |
Mueller B (2015) Experimental interactions between clay minerals and bacteria: a review. Pedosphere 25: 799–810. doi: 10.1016/S1002-0160(15)30061-8
|
| [43] |
Kostka JE, Dalton DD, Skelton H, et al. (2002) Growth of iron (Ⅲ)-reducing bacteria on clay minerals as the sole electron acceptor and comparison of growth yields on a variety of oxidized iron forms. Appl Environ Microbiol 68: 6256–6262. doi: 10.1128/AEM.68.12.6256-6262.2002
|
| [44] | Ahmed Z, Ivanov V, Hyun SH, et al. (2001) Effect of divalent iron on methanogenic fermentation of fat-containing wastewater. Environ Engrg Res 6:139–146. |
| [45] |
Li Z, Wrenn BA, Venosa AD (2006) Effects of ferric hydroxide on methanogenesis from lipids and long-chain fatty acids in anaerobic digestion. Water Environ Res 78: 522–530. doi: 10.2175/106143005X73064
|
| [46] |
Ivanov V, Stabnikova EV, Stabnikov VP, et al. (2002) Effects of iron compounds on the treatment of fat-containing wastewaters. Appl Biochem Micro 38: 255–258. doi: 10.1023/A:1015475425566
|
| [47] |
Bampalioutas K, Vlysidis A, Lyberatos G, et al. (2019) Detoxification and methane production kinetics from three-phase olive mill wastewater using Fenton's reagent followed by anaerobic digestion. J Chem Technol Biot 94: 265–275. doi: 10.1002/jctb.5772
|
| [48] |
Baek G, Kim J, Shin SG, et al. (2016) Bioaugmentation of anaerobic sludge digestion with iron-reducing bacteria: process and microbial responses to variations in hydraulic retention time. Appl Microbiol Biot 100: 927–937. doi: 10.1007/s00253-015-7018-y
|
| [49] |
Park CM, Novak JT (2013) The effect of direct addition of iron(Ⅲ) on anaerobic digestion efficiency and odor causing compounds. Water Sci Technol 68: 2391–2396. doi: 10.2166/wst.2013.507
|
| [50] |
Yue ZB, Ma D, Wang J, et al. (2015) Goethite promoted anaerobic digestion of algal biomass in continuous stirring-tank reactors. Fuel 159: 883–886. doi: 10.1016/j.fuel.2015.07.059
|
| [51] | Capson-Tojo G, Girard C, Rouez M, et al. (2018) Addition of biochar and trace elements in the form of industrial FeCl3 to stabilize anaerobic digestion of food waste: dosage optimization and long-term study. J Chem Technol Biot 94: 505–515. |
| [52] |
García-Balboa C, Cautivo D, Blázque, ML, et al. (2010) Successive ferric and sulphate reduction using dissimilatory bacterial cultures. Water Air Soil Poll 207: 213–226. doi: 10.1007/s11270-009-0130-9
|
| [53] |
Wang MW, Zhao Z, Zhang Y (2018) Sustainable strategy for enhancing anaerobic digestion of waste activated sludge: driving dissimilatory iron reduction with Fenton sludge. ACS Sustain Chem Eng 6: 2220–2230. doi: 10.1021/acssuschemeng.7b03637
|
| [54] |
Flores-Alsina X, Solon K, Mbamba CK, et al. (2016) Modelling phosphorus (P), sulfur (S) and iron (Fe) interactions for dynamic simulations of anaerobic digestion processes. Water Res 95: 370–382. doi: 10.1016/j.watres.2016.03.012
|
| [55] |
Yap SD, Astals S, Lu Y, et al. (2018) Humic acid inhibition of hydrolysis and methanogenesis with different anaerobic inocula. Waste Manage 80: 130–136. doi: 10.1016/j.wasman.2018.09.001
|
| [56] |
Khadem AF, Azman S, Plugge CM, et al. (2017) Effect of humic acids on the activity of pure and mixed methanogenic cultures. Biomass Bioenerg 99: 21–30. doi: 10.1016/j.biombioe.2017.02.012
|
| [57] |
Stepanov N, Senko O, Perminova I, et al. (2019) A new approach to assess the effect of various humic compounds on the metabolic activity of cells participating in methanogenesis. Sustainability 11: 3158. doi: 10.3390/su11113158
|
| [58] |
Greenland DJ (1971) Interactions between humic and fulvic acids and clays. Soil Sci 111: 34–41. doi: 10.1097/00010694-197101000-00004
|
| [59] | Boguta P, D'Orazio V, Senesi N, et al. (2019) Insight into the interaction mechanism of iron ions with soil humic acids. The effect of the pH and chemical properties of humic acids. J Environ Manage 245: 367–374. |
| [60] | Tay JH, Tay STL, Ivanov V, et al. (2008) Compositions and methods for the treatment of wastewater and other waste. US Patent 7393452. |
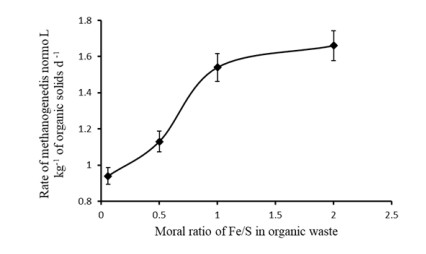
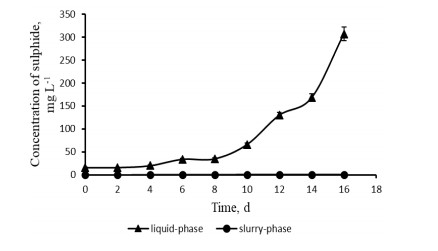
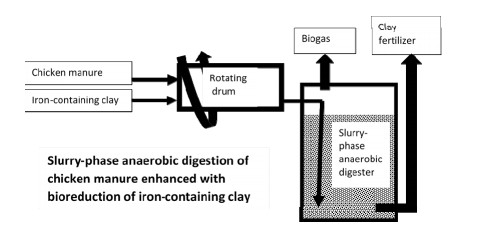
Figures(3) / Tables(2)

Volodymyr Ivanov, Viktor Stabnikov, Olena Stabnikova, Anatoliy Salyuk, Evhenii Shapovalov, Zubair Ahmed, Joo Hwa Tay. Iron-containing clay and hematite iron ore in slurry-phase anaerobic digestion of chicken manure[J]. AIMS Materials Science, 2019, 6(5): 821-832. doi: 10.3934/matersci.2019.5.821







 DownLoad:
DownLoad: