Citation: Sasha Tozzi, Walker O. Smith. Contrasting Photo-physiological Responses of the Haptophyte Phaeocystis Antarctica and the Diatom Pseudonitzschia sp. in the Ross Sea (Antarctica)[J]. AIMS Geosciences, 2017, 3(2): 142-162. doi: 10.3934/geosci.2017.2.142

| [1] | Kirk JTO (1994) Light and photosynthesis in aquatic ecosystems. Cambridge University Press, Cambridge, UK. |
| [2] |
Jones RM and Smith WO Jr (2017) The influence of short-term events on the hydrographic and biological structure of the southwestern Ross Sea. J Mar Systems 166: 184-195. doi: 10.1016/j.jmarsys.2016.09.006

|
| [3] |
Smith WO Jr and Nelson DM (1985) Phytoplankton bloom produced by a receding ice edge in the Ross Sea: spatial coherence with the dendity field. Sci 227: 163-166. doi: 10.1126/science.227.4683.163
|
| [4] |
Arrigo KR and McClain CR (1994) Spring phytoplankton production in the western Ross Sea. Sci 266: 261-263. doi: 10.1126/science.266.5183.261
|
| [5] |
Arrigo KR, Robinson DH, Worthen DL, et al. (1999) Phytoplankton community structure and the drawdown of nutrients and CO2 in the Southern Ocean. Sci 283: 365-367. doi: 10.1126/science.283.5400.365
|
| [6] |
Smith WO Jr and Gordon LI (1997) Hyperproductivity of the Ross Sea (Antarctica) polynya during austral spring. Geophys Res Letters 24: 233-236. doi: 10.1029/96GL03926
|
| [7] |
Rousseau V, Chrétiennot-Dinet M-J, Jacobsen A, et al. (2007) The life cycle of Phaeocystis: state of knowledge and presumptive role in ecology. Biogeochem 83: 29-47. doi: 10.1007/s10533-007-9085-3
|
| [8] |
Smith WO Jr, Ainley DG, Arrigo KR, et al (2014) The oceanography and ecology of the Ross Sea. Annu Rev Mar Sci 6: 469-487. doi: 10.1146/annurev-marine-010213-135114
|
| [9] |
Smith WO Jr and Jones RM (2015) Vertical mixing, critical depths, and phytoplankton growth in the Ross Sea. ICES J. Mar. Sci 72: 1952-1960. doi: 10.1093/icesjms/fsu234
|
| [10] |
Leventer A and Dunbar RB (1996) Factors influencing the distribution of diatoms and other algae in the Ross Sea. J Geophys Res 101: 18489-18500. doi: 10.1029/96JC00204
|
| [11] | Nuccio C, Innamorati M, Lazzara L, et al. (2000) Spatial and temporal distribution of phytoplankton coenoses in the Ross Sea. In: Faranda FM, Guglielmo L, Ianora A (Eds.), Ross Sea Ecology. Italiantartide Expeditions (1987-1995). Springer Verlag, Berlin, 231-245. |
| [12] | Fonda Umani S, Accornero A, Budillon G, et al(2002) Particulate matter and plankton dynamics in the Ross Sea Polynya of Terra Nova Bay during the austral aummer 1997/98. J Mar Syst 36: 29-49. |
| [13] | McGillicuddy DM Jr, Sedwick PN, Dinniman MS, et al. (2015) Iron supply and demand in an Antarctic shelf system. Geophys Res Letters 42:. |
| [14] | Falkowski PG and Raven JA (1997) Aquatic Photosynthesis. Princeton Univ Press, Princeton, NJ. |
| [15] |
Kropuenske LR, Mills MM, van Dijken GL, et al. (2009) Photophysiology in two major Southern Ocean phytoplankton taxa: Photoprotection in Phaeocystis antarctica and Fragilariopsis cylindrus. Limnol Oceanogr 54: 1176-1196. doi: 10.4319/lo.2009.54.4.1176
|
| [16] | Vass I, Styring S, Hundal T, et al (1992) Reversible and irreversible intermediates during photoinhibition of photosystem II: stable reduced QA species promote chlorophyll triplet formation. Proc Nat Acad Sci US 89 1408-1412. |
| [17] |
Demmig-Adams B and Adams WW (1992) Photoprotection and other responses of plants to high light stress. Annu Rev Plant Physiol Plant Mol Biol 43: 599-626. doi: 10.1146/annurev.pp.43.060192.003123
|
| [18] | Smith WO Jr and Donaldson K (2015) Photosynthesis-irradiance responses in the Ross Sea, Antarctica: a meta-analysis. Biogeosci 12: 1-11. |
| [19] |
Arrigo KR, Mills MM, Kropuenske LR, et al (2010) Photophysiology in two major southern ocean phytoplankton taxa: photosynthesis and growth of Phaeocystis antarctica and Fragilariopsis cylindrus under different irradiance levels. Integr Compar Biol 50: 950–966. doi: 10.1093/icb/icq021
|
| [20] |
Smith WO Jr, Tozzi S, Sedwick PW, et al. (2013) Spatial and temporal variations in variable fluorescence in the Ross Sea (Antarctica): environmental and biological correlates. Deep-Sea Res I 79: 141-155. doi: 10.1016/j.dsr.2013.05.002
|
| [21] |
Genty B, Briantais JM, Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990: 87-92. doi: 10.1016/S0304-4165(89)80016-9
|
| [22] |
Kolber ZS, Prasil O, Falkowski PG (1998) Measurements of variable chlorophyll fluorescence using fast repetition rate techniques: defining methodology and experimental protocols. Biochim Biophys Acta 1367: 88-106. doi: 10.1016/S0005-2728(98)00135-2
|
| [23] | Keller MD, Selvin RC, Claus W, et al. (1987) Medifor the culture of oceanoic ultraphytoplankton. J Phycol 23: 633-638. |
| [24] | Sedwick PN, Marsay CM, Aguilar-Islas AM, et al. (2011) Early-season depletion of dissolved iron in the Ross Sea polynya: implications for iron dynamics on the Antarctic continental shelf. J Geophys Res 116. |
| [25] |
Kustka AB, Jones BM, Hatta M, et al. (2015) The influence of iron and siderophores on eukaryotic phytoplankton growth rates and community composition in the Ross Sea. Mar Chem 173: 195-207. doi: 10.1016/j.marchem.2014.12.002
|
| [26] | Ryan-Keogh TJ, Delizo LM, Smith WO Jr, et al. (2016) Temporal progression of photosynthetic strategy by phytoplankton in the Ross Sea, Antarctica. J Mar Systems 166: 87-96. |
| [27] |
Moisan TA, Mitchell BG (1999) Photophysiological acclimation of Phaeocystis antarctica Karsten under light limitation. Limnol Oceanogr 44: 247-258. doi: 10.4319/lo.1999.44.2.0247
|
| [28] |
Moisan TA, Olaziola M, Mitchell BG (1998) Xanthophyll cycling in Phaeocystis antarctica: changes in cellular fluorescence. Mar Ecol Prog Ser 169: 113-121. doi: 10.3354/meps169113
|
| [29] | Van Leuwe MA and Stefels J (1998) Effects of iron and light stress on the biochemical composition of Antarctic Phaeocystis sp.(Prymnesiophyceae). II. Pigment composition. J Phycol 34: 496-503. |
| [30] |
Liselotte R and Dale R. (1997) Photoinduction of UV-absorbing compounds in Antarctic diatoms and Phaeocystis antarctica. Mar Ecol Prog Ser 160: 13-25. doi: 10.3354/meps160013
|
| [31] | Tang KW, Smith WO Jr, Shields AR, et al. (2009) Survival and recovery of Phaeocystis antarctica (Prymnesiophyceae) from prolonged darkness and freezing. Phil Trans Roy Soc, serB 276: 81-90. |
| [32] |
Caron DA, Dennett MR, Lonsdale DJ, et al. (2000) Micro-zooplankton herbivory in the Ross Sea, Antarctica. Deep Sea Res II 47: 3249-3272. doi: 10.1016/S0967-0645(00)00067-9
|
| [33] |
Bertrand EM, Saito MA, Rose JM, et al. (2007) Vitamin B12 and iron co-limitation of phytoplankton growth in the Ross Sea. Limnol Oceanogr 52: 1079-1093. doi: 10.4319/lo.2007.52.3.1079
|
| [34] |
Sedick PN and DiTullio GR (1997) Regulation of algal blooms in Antarctic shelf waters by the release of iron from melting sea ice. Geophys Res Letters 24: 2515-2518. doi: 10.1029/97GL02596
|
| [35] |
Feng Y, Hare CE, Rose JM, et al. (2010) Interactive effects of CO2, irradiance and iron on Ross Sea phytoplankton. Deep Sea Res I 57: 368-383. doi: 10.1016/j.dsr.2009.10.013
|
| [36] |
Rose JM, Feng Y, DiTullio G, et al. (2009) Synergistic effects of iron and temperature on Antarctic plankton assemblages. Biogeosci 6: 3131–3147. doi: 10.5194/bg-6-3131-2009
|
| [37] |
Xu K, Fu F-X, Hutchins DA (2014) Comparative responses of two dominant Antarctic phytoplankton taxa to interactions between ocean acidification, warming, irradiance, and iron availability. Limnol Oceanogr 59: 1919-1931. doi: 10.4319/lo.2014.59.6.1919
|
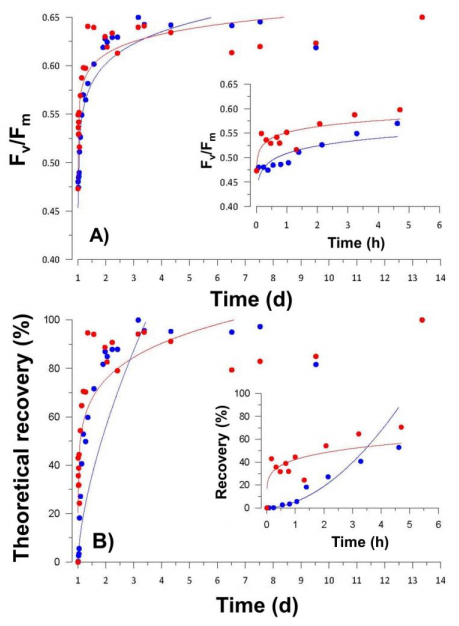
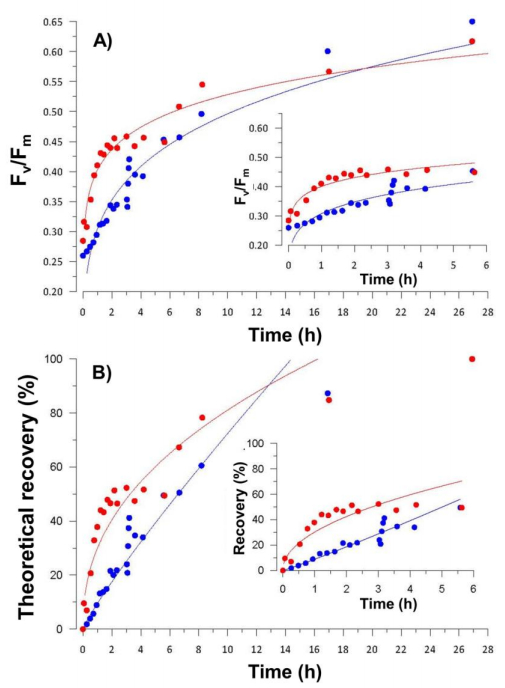
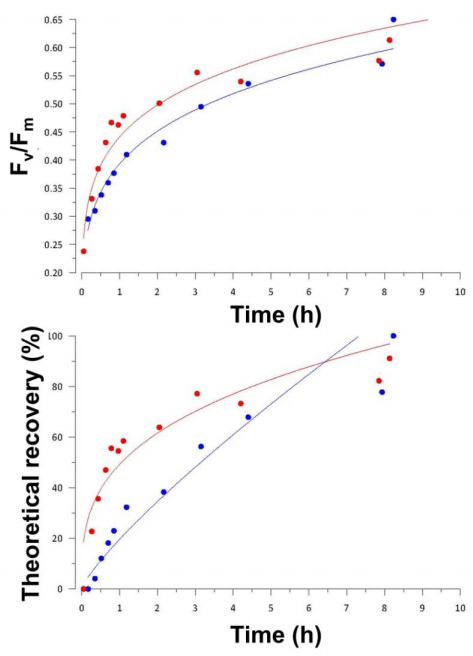
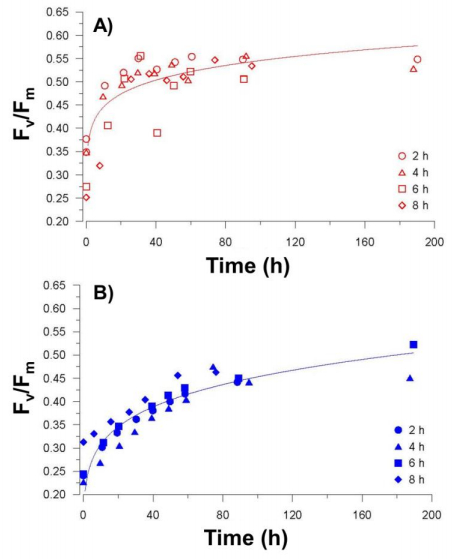
Figures(7) / Tables(2)

Sasha Tozzi, Walker O. Smith. Contrasting Photo-physiological Responses of the Haptophyte Phaeocystis Antarctica and the Diatom Pseudonitzschia sp. in the Ross Sea (Antarctica)[J]. AIMS Geosciences, 2017, 3(2): 142-162. doi: 10.3934/geosci.2017.2.142







 DownLoad:
DownLoad: